



Bolitaeninae
Richard E. Young




This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
These are common meso- to bathypelagic octopods of rather small size (ca. 85 mm ML in the largest species) that are broadly distributed mostly in tropical to temperate latitudes of all oceans. They have gelatinous bodies, numerous chromatophores and possess short arms with a single series of suckers. The mantle aperture is wide. The hectocotylus has a swollen ligula in Bolitaena but is unknown in Japetella.
Brief diagnosis:
An incirrate octopod ...
- with ctenoglossan radula.
- with laterally compressed eyes.
Characteristics
- Arms
- Arms short; length less than ML.
- Suckers in a single series.
- Third right arm in Bolitaena hectocotylized (Japetella apparently lacks hectocotylization although some sexual dimorphism of arms III occurs).
- Eyes
- Eyes laterally compressed.
- Eyes laterally compressed.
- Viscera
- Long axis of digestive gland parallels body axis.
- Stomach posterior to digestive gland.
- Radula
- Radula with multicuspid lateral teeth (= ctenoglossan radula).
Click on an image to view larger version & data in a new window
Figure. Radula of Bolitaena pygmaea, redrawn from Thore, 1949.
- Photophores
- Mature females with a circular light organ surrounding the mouth.
Comments
For immature octopods, the two genera can be difficult to separate. The most obvious features are the size of the eyes (larger in Japetella) and the distance between the eyes (larger in Bolitaena). When comparing adjacent octopods (see below), these differences are distinct. However, with isolated specimens, the differences are not always obvious.

Figure. Left - Dorsal view of Japetella diaphana, 83 mm ML, nearly mature female. Right - Dorsal view of Bolitaena pygmaea, 38 mm ML, immature female. Differences above in arm sizes and pigmentation is accuated by the size disparity. In addition, if the J. diaphana were smaller, the separation of its eyes would be less.
Nomenclature
Bolitaena microcotyla Steenstrup in Hoyle, 1886, has many similarities to Eledonella pygmaea. Thore (1949), in his review of the family, distinguishes B.microcotyla primarily by its having (1) large eggs, (2) suckers apart within the web and closely set beyond the web and (3) reddish-purple color. The eggs of B. microcotyla are 2.5 mm in length (Thore, 1949). The eggs of "E. pygmaea" (incorrectly identified by Young, 1972, as Bolitaena microcotyla), however, are nearly the same length (2.0 mm, Young, 1972). The second and third distinguishing characters result from rather marked morphological changes that bolitaenids exhibit at sexual maturity. At maturity females lose the irridescence on their eyes and digestive glands, become heavily pigmented (the degree of pigmentation is greater in Bolitaena), and the arms increase in length relative to the mantle length. The longer arms produce the distinctive sucker arrangement on the arms (Note the pigmentation and sucker spacing in the photograph of the brooding female below). Unfortunately, adequate material has not been available to quantitatively document these changes. Nevertheless, the mature female appears very different from the subadult and has resulted in labeling different life-history stages as different genera.
The genus Bolitaena was described nearly 30 years before a species name, B. microcotyla, was appended in 1886, in a brief description. Description of a new genus without a type species is no longer legal under the rules of zoological nomenclature but was valid at the time. Eledonella pygmaea was described by Verrill in 1884. The proper name, therefore, is Bolitaena pygmaea (Verrill, 1884).
Careful comparisons of specimens in good condition (or, more likely genetic analyses) may eventually reveal that more than one species of Bolitaena exists. Our present knowledge, however, requires synonymizing the genera and leaves little reason for maintaining separate species.
Thore (1949) described Dorsopsis taningi as a new genus and species of bolitaenid that had hemispherical, dorsally directed eyes. This specimen, however, was a damanged Japetella (Hochberg, et al., 1992).
A list of all nominal genera and species in the Bolitaeninae can be found here. The list includes the current status and type species of all genera, and the current status, type repository and type locality of all species and all pertinent references.
Life history
A large light organ develops around the mouth in females at maturity. Presumably it functions in attracting a male in dark waters at a depth of 1000m or more (Robison and Young, 1981). This is the only luminescent organ known in incirrate octopods.

Figure. Oral view of mature female Japetella diaphana showing yellow photophore around mouth, off Hawaii. Photograph by M. Vecchione.
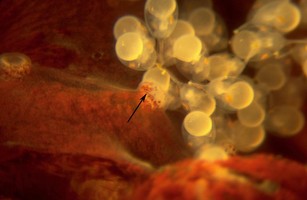
Figure. Arms of Bolitaena pygmaea holding onto a brood of embryos. Arrow in photograph points to a sucker holding one of the eggs. Photograph by R. Young.
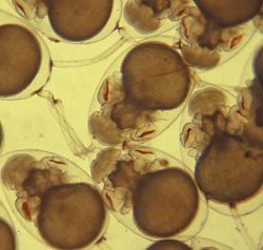
Figure. Developing embryos of Bolitaena pygmaea or Japetella diaphana. The egg in the upper left corner of the photograph has two stalks attached to the chorion opposite the large yolk sac; the lower of these two stalks has a third stalk attached to it (note the pigmentation that marks the attachment). Photograph by R. Young.
Off Hawaii female bolitaenids apparently release their young at a depth near 800m. The young, free-swimming octopods are found at depths around 150 - 250 m. As they grow they descend, at first abruptly then more gradually, to depths of 800 -1400 m. Gravid females are found at the lower end of the range where mating presumably occurs (Young, 1978). More details of this pattern are presented under each genus.
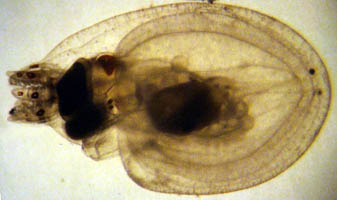
Figure. Ventrolateral view of a hatchling of Japetella diaphana. This hatchling may have hatched slightly prematurely due to the trauma of capture). Note that only a few chromatophores are present and that the arms are very short. Photograph by R. Young.
References
Hochberg, F. G., M. Nixon and R. B. Toll. 1992. Order Octopoda Leach, 1818. In: Sweeney, M. J., C. F. E. Roper, K. M. Mangold, M. R. Clarke and S. v. Boletzky (eds.) "Larval" and juvenile cephalopods: A manual for their identification. Smithson. Contr. Zool., 513:1-282.
Robison, B. and R.E. Young. 1981. Bioluminescence in pelagic octopods. Pac. Sci., 35:39-44.
Thore, S. 1949. Investigations on the "Dana" Octopoda. Dana-Report No. 33: 1-85.
Young, R. E. 1978. Vertical distribution and photosensitive vesicles of pelagic cephalopods from Hawaiian waters. Fish. Bull. 76: 583-615.
Title Illustrations

| Scientific Name | Japetella diaphana |
|---|---|
| Location | off Hawaii |
| Specimen Condition | Live Specimen |
| View | Side |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 1995

|
About This Page
University of Hawaii, Honolulu, HI, USA
Page copyright © 2018
 Page: Tree of Life
Bolitaeninae .
Authored by
Richard E. Young.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Bolitaeninae .
Authored by
Richard E. Young.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 10 October 2017
Citing this page:
Young, Richard E. 2017. Bolitaeninae . Version 10 October 2017 (under construction). http://tolweb.org/Bolitaeninae/20193/2017.10.10 in The Tree of Life Web Project, http://tolweb.org/

