



Dissorophoidea 
Adam Huttenlocker


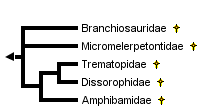


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Dissorophoids comprise a diverse group of small and medium-sized temnospondyl stegocephalians that occupied a number of ecological niches (both aquatic and terrestrial) and played an important role in vertebrate communities for at least 80 million years (Carroll, 1964; Holmes, 2000). Some of the earliest known dissorophoid fossils occur in Pennsylvanian-aged sediments (325-299 million years ago) of the Czech Republic and the Midwestern United States. They are represented by the tiny, paedomorphic Branchiosauridae and Micromelerpetontidae. These forms were virtually ubiquitous in aquatic freshwater vertebrate communities of the Pennsylvanian and Early Permian in Europe and North America. Both groups had aquatic larvae and adult branchiosaurids seem to have retained an ossified hyobranchial skeleton into maturity, along with external gill filaments and gill rakers (Holmes, 2000). These taxa resided in a niche similar to that which is occupied by some ambystomatid salamanders (e.g., the Mexican axolotl) today.
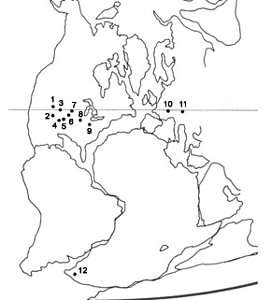
Figure 1. Paleobiogeographic distribution of Permo-Carboniferous and Triassic dissorophoids based on selected fossil localities. (1) Utah, USA (Lower Permian); (2) Rio Arriba County, north-central New Mexico, USA (Lower Permian); (3) Badger Creek quarry, central Colorado, USA (Upper Pennsylvanian); (4) Baylor County localities, Texas, USA (Lower Permian); (5) Fort Sill fissure fill, Oklahoma, USA (Lower Permian); (6) Hamilton quarry, Kansas, USA (Upper Pennsylvanian); (7) Mayer Farm locality, Humboldt, Nebraska, USA (Lower Permian); (8) Mazon Creek, Illinois, USA (Upper Pennsylvanian); (9) Linton Coal, Ohio, USA (Upper Pennsylvanian); (10) Rotliegendes, Germany (Upper Pennsylvanian / Lower Permian); (11) Nýřany, Czech Republic (Upper Pennsylvanian); (12) Micropholis localities, Karoo Basin, South Africa (Lower Triassic). (Modified from Kissel and Reisz, 2004.) © Adam Huttenlocker
Another taxon, the Amphibamidae, first appears in slightly younger deposits than the earliest dissorophoids, ranging from the Late Pennsylvanian to the Early Triassic. It was composed of miniaturized temnospondyls showing predominantly terrestrial adaptations based on the development and degree of ossification of the appendicular skeleton and histological evidence in the long bones of some specimens (Bolt, 1977, Laurin et al., 2004). While these adaptations are most evident in adults, amphibamids still relied on water for reproduction and had aquatic larvae with external gills, closely resembling branchiosaurs (Milner, 1982). Amphibamidae was the longest-lived dissorophoid taxon, persisting for at least 60 million years.
The latest dissorophoid clades that appear in the fossil record are the trematopids (Late Pennsylvanian to Early Permian) and the armored dissorophids (Early to Late Permian). The trematopids are typified by a posteriorly elongated or keyhole-shaped external naris (as in Ecolsonia) and large ‘caniniform’ maxillary teeth. The most widely recognized feature of the Dissorophidae, on the other hand, is the development of an armored carapace covering the dorsal surface of the body. Together, these two taxa display more progressive trends in terrestriality, larger adult body sizes, and an otic notch that is often closed off posteriorly.
Characteristics
The skulls of some selected dissorophoids are illustrated in Figure 2 for a comparison of the major cranial features, particularly in the region of the otic notch. In general, the antorbital region of the skull is very short and broad with large, widely spaced nares, and the posterior region of the skull is usually abbreviated when compared to other early stegocephalians (particularly in branchiosaurids and most amphibamids where the tabular and postparietal form short, transverse bars). The otic notch, believed by some authors to be associated with the reception of airborne sounds (Bolt, 1974; Holmes, 2000), is also an important feature in dissorophoids. However, the role of the otic notch for hearing air-borne sounds has recently been disputed (Laurin, 1998; Laurin and Soler-Gijon, 2006). Among Permo-Carboniferous temnospondyls, the otic notch is most deeply developed in this group, particularly in the trematopids and dissorophids where it is quite long anteroposteriorly and sometimes closed off posteriorly (fig. 2C,D).
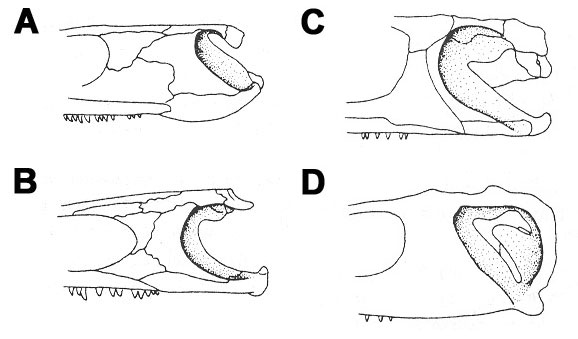
Figure 2. Evolutionary trends in the development of the otic notch based on selected temnospondyls. Posterior region of the skull (not to scale) is shown in left lateral view. (A) edopoid Dendrerpeton; (B) amphibamid dissorophoid Eoscopus; (C) dissorophid Dissorophus; (D) cacopine dissorophid Cacops. (modified from Holmes et al., 1998). © Adam Huttenlocker.
A complete list of synapomorphies supporting the monophyly of Dissorophoidea is listed below (from Daly, 1994; Schoch and Rubidge, 2005):
- Laterally exposed palatine (absent or reduced in some branchiosaurs and amphibamids)
- Ventral prefrontal process contacts dorsal process of palatine medial to the lacrimal within the orbit (absent in some amphibamids)
- Dorsal quadrate process
- Posteromedial process of the quadratojugal
- Supratympanic flange on dorsal border of the otic notch in some dissorophoids (absent in branchiosaurids, micromelerpetontids, and some amphibamids)
- Internal nasal flange, located ventrally in the snout
- Pterygoid-vomer contact lost (except in trematopids)
- Interclavicle anterior process reduced
- Contribution of frontals to dorsal orbital margins (except in some amphibamids: Platyrhinops and Amphibamus)
Discussion of Phylogenetic Relationships
The phylogenetic position of Dissorophoidea within Temnospondyli is generally well resolved, with most authors favoring a sister-group relationship to eryopoids or zatrachydid temnospondyls (collectively the “Euskelia” of Yates and Warren, 2000). It is very likely that the earliest members of the group evolved from a small, eryopoid or zatrachydid-like ancestor. Within Dissorophoidea, five major taxa, usually recognized as families, are currently recognized, including the Branchiosauridae, Micromelerpetontidae, Amphibamidae, Trematopidae, and Dissorophidae (discussed above). It is beyond this point that the interrelationships of the various dissorophoid families are less agreed upon.
An early cladistic analysis performed by Trueb and Cloutier (1991) supported a monophyletic Amphibamidae (= ‘Doleserpetontidae’), uniting the taxa Tersomius, Doleserpeton, and Amphibamus within the group and suggesting close affinities to the Dissorophidae. Daly (1994) provided the first substantial cladistic analysis of the relationships of dissorophoids, but did not explore genus-level relationships in any detail and assumed the monophyly of most of the known dissorophoid families. She suggested that Dissorophidae was monophyletic and represented the sister taxon to Trematopidae, and that Amphibamidae was more closely related to dissorophids and trematopids than to the more basal branchiosaurids. Yates and Warren (2000) performed an analysis of temnospondyls, demonstrating a monophyletic Euskelia (discussed above). The authors further demonstrated relationships within Dissorophoidea consistent with the hypothesis of Daly (1994), supporting Dissorophidae and Trematopidae as the monophyletic sister group to Amphibamidae (represented by the genus Tersomius).
More recently, Ruta, Jeffery, and Coates (2003) demonstrated Amphibamidae as the monophyletic sister taxon to Branchiosauridae or to Branchiosauridae + Crown-Lissamphibia (including modern amphibians), with Trematopidae and Dissorophidae serving as successive outgroups. Their phylogenetic hypothesis suggests a basal position for trematopids within Dissorophoidea. Other recent analyses (Anderson, in press; Anderson et al., 2004) have indicated similar familial-level relationships, suggesting close affinities between amphibamids, ‘branchiosaurs,’ and at least some lissamphibians, and placing Trematopidae in a relatively basal position. However, the most recent analyses excluded the family Dissorophidae and other important dissorophoid taxa. Given these recent views, a better consensus of dissorophoid relationships will have to await systematic revisions of key genera and more detailed anatomical descriptions of well preserved specimens (Anderson and Bolt, 2005).
References
Anderson, J. S. In press. Incorporating ontogeny into the matrix: a phylogenetic evaluation of developmental evidence for the origin of modern amphibians; pp. XX-XX in J. Anderson and H.-D. Sues (eds.), Major Transitions in Vertebrate Evolution. Indiana University Press, Bloomington.
Anderson, J. S. and J. R. Bolt. 2005. A nearly complete amphibamid skull from Richards Spur (Fort Sill), Oklahoma, and its implications for interpretation of dental characters related to the origin of modern amphibians. Journal of Vertebrate Paleontology 25(3, supplement):31A.
Anderson, J. S., S. Sumida, D. Berman, A. Henrici, and T. Martens. 2004. The dissorophoid temnospondyls from the Early Permian of Bromacher, Germany. Journal of Vertebrate Paleontology 24(3, supplement):34A.
Bolt, J. R. 1974. A trematopsid skull from the Lower Permian, and analysis of some characters of the dissorophoid (Amphibia: Labyrinthodontia) otic notch. Fieldiana Geology 30(3):67-79.
Bolt, J. R. 1977. Dissorophoid relationships and ontogeny, and the origin of the Lissamphibia. Journal of Paleontology 51:235-249.
Boy, J. A. and H.-D. Sues. 2000. Branchiosaurs: larvae, metamorphosis and heterochrony in temnospondyls and seymouriamorphs; Pp. 1150-1197 in H. Heatwole and R. L. Carroll (eds.) Amphibian Biology, Volume 4. Surrey Beatty and Sons Ltd., Australia.
Carroll, R. L. 1964. Early evolution of the dissorophid amphibians. Bulletin of the Museum of Comparative Zoology, Harvard University 131(7):161-250.
Daly, E. 1994. The Amphibamidae (Amphibia: Temnospondyli), with a description of a new genus from the Upper Pennsylvanian of Kansas. University of Kansas Museum of Natural History, Miscellaneous Publication No. 85: 59 pp.
DeMar, R. E. 1968. The Permian labyrinthodont amphibian Dissorophus multicinctus, and adaptations and phylogeny of the family Dissorophidae. Journal of Paleontology 42:1210-1242.
Dilkes, D. W. 1990. A new trematopsid amphibian (Temnospondyli: Dissorophoidea) from the Lower Permian of Texas. Journal of Vertebrate Paleontology 10(2):222-243.
Holmes, R. 2000. Palaeozoic temnospondyls; Pp. 1081-1120 in H. Heatwole and R. L. Carroll (eds.) Amphibian Biology, Volume 4. Surrey Beatty and Sons Ltd., Australia.
Holmes, R. B., R. L. Carroll, and R. R. Reisz. 1998. The first articulated skeleton of Dendrerpeton acadianum (Temnospondyli, Dendrerpetontidae) from the Lower Pennsylvanian locality of Joggins, Nova Scotia, and a review of its relationships. Journal of Vertebrate Paleontology 18(1):64-79.
Kissel, R. A., and R. R. Reisz. 2004. Synapsid fauna of the Upper Rock Lake Shale near Garnett, Kansas and the diversity pattern of early amniotes; Pp.409-428 in G. Arratia, M. V. H. Wilson, and R. Cloutier (eds.) Recent Advances in the Origin and Early Radiation of Vertebrates. Verlag Dr. Friedrich Pfeil, München.
Laurin, M. 1998. The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I-systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: 1-42.
Laurin, M., and R. Soler-Gijon. 2006. The oldest known stegocephalian (Sarcopterygii: Temnospondyli) from Spain. Journal of Vertebrate Paleontology 26(2):284-299.
Laurin, M., M. Girondot, and M.-M. Loth. 2004. The evolution of long bone microstructure and lifestyle in lissamphibians. Paleobiology 30(4):589-613.
Lewis, G. E. and P.P. Vaughn. 1965. Early Permian vertebrates from the Cutler Formation of the Placerville area, Colorado. U. S. Geological Survey Professional Papers 503C.
Milner, A. R. 1982. Small temnospondyl amphibians from the middle Pennsylvanian of Illinois. Palaeontology 25(3):635-664.
Ruta, M., J. E. Jeffery, and M. I. Coates. 2003. A supertree of early tetrapods. Proceedings of the Royal Society of London Series B 270:2507-2516.
Schoch, R. R. and B. S. Rubidge. 2005. The amphibamid Micropholis from the Lystrosaurus Assemblage Zone of South Africa. Journal of Vertebrate Paleontology 25:502-522.
Trueb, L. and R. Cloutier. 1991. A phylogenetic investigation of the inter- and intrarelationships of the Lissamphibia (Amphibia: Temnospondyli); pp. 223-313 in H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus. Cornell University Press, Ithaca.
Yates, A. M. and A. A. Warren. 2000. The phylogeny of the higher temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli. Zoological Journal of the Linnean Society 128:77-121.
Title Illustrations

| Scientific Name | Ecolsonia cutlerensis |
|---|---|
| Location | North-central New Mexico |
| Specimen Condition | Fossil -- Period: Early Permian (Cutler Fm) |
| Identified By | Berman et al., 1985 |
| Copyright |
© Adam Huttenlocker

|
About This Page
Adam Huttenlocker
California State University, San Bernardino, California, USA
Correspondence regarding this page should be directed to Adam Huttenlocker at
Page copyright © 2009 Adam Huttenlocker
All Rights Reserved.
- First online 12 March 2007
- Content changed 12 March 2007
Citing this page:
Huttenlocker, Adam. 2007. Dissorophoidea . Version 12 March 2007 (under construction). http://tolweb.org/Dissorophoidea/17607/2007.03.12 in The Tree of Life Web Project, http://tolweb.org/

