



Lophiidae
Goosefishes, monkfishes
John H. Caruso
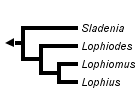


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The monkfishes are a family of anglerfishes that includes 25 species distributed among four genera (Sladenia, Lophiodes, Lophiomus, and Lophius). These fishes inhabit soft bottoms of the continental slope, usually at depths beyond 200 m, with some species inhabiting depths in excess of 1000 m. Juveniles are often captured at mesopelagic depths. Some species of the genus Lophius can grow to over 1 m and are often exploited as highly esteemed food fishes.
Characteristics
Diagnosis
Lophiiform fishes with thin, loose, scale-less skin bearing well-developed fleshy tendrils or cirri on lateral margins of head, lower jaw, and body. Head and body greatly depressed. Mouth wide, cavernous, upper and lower jaws bearing long, slender, depressible teeth; lower jaw projecting well beyond upper. Vomer, palatine, ceratobranchial V, and pharyngobranchials II-IV dentigerous. Gill filaments present on first three branchial arches, absent on fourth. Dorsal surface of head with numerous bony knobs, spines, and ridges; frontal bone with well-developed supraorbital ridge, bearing two to four, dorsolaterally directed spines; a frontal ridge running forward from supraorbital ridge to tip of snout; one to three sphenotic spines; a single pterotic spine; palatine with two dorsally directed spines at tip of snout; interopercle with one to three spines at posterodorsal angle. Gill openings extending below and behind or below, behind, and in front of bases of pectoral fin. Spinous dorsal fin primitively of six spines, divided into cephalic (two or three spines) and post-cephalic portions (one to three spines); illicium and second dorsal-fin spine inserted close together on common pterygiophore. Pelvic fins present.
Coloration uniform gray, brown to dark brown, somewhat lighter on belly; usually without markings of any kind (dorsal surface of head and body mottled in Sladenia).
Discussion of Phylogenetic Relationships

© Christopher P. Kenaley
The relationships of the Lophiidae as presented by Caruso (1985). For character states, click here: 1-6, 7-12, 13-15, 16-17, 18, and 19.
The following 19 characters were used in a phylogenetic analysis of the family Lophiidae by Caruso (1985).
- Body shape. A rounded robust body shape is characteristic of Sladenia, most other lophiiforms (ogcocephalids and the ceratioid genus Thaumatichthys are the only exceptions) and all other paracanthopterygian fishes. Lophiodes exhibits a depressed head and body; Lophiomus and Lophius are very strongly depressed. Outgroup comparison indicates that the depressed body shape is the derived state of this character.
- Anterior articular spines. The articular bone is devoid of spines in Sladenia and in other lophiiforms and paracanthopterygians. The remaining lophiid genera have a single spine anterior to the jaw joint; Lophiodes has an additional spine posterior to the jaw joint. The presence of articular spines is thus the derived state of this character.
- Quadrate spines. The quadrate bone is devoid of spines in Sladenia and in other lophiiforms and paracanthopterygians. The remaining genera have a single prominent spine extending outward from the dorsolateral edge of the articulating surface of the jaw joint. In Lophius an additional spine is present dorsomedial to the aforementioned spine. The presence of quadrate spines is thus derived.
- Humeral spines. The humeral spine is a large complex spine that extends posteriorly from the posterior end of the vertical limb of the cleithrum in Lophiodes, Lophiomus, and Lophius. It is absent in Sladenia, other lophiiforms, and other paracanthopterygians. Its presence is thus derived.
- Subopercular spines. The subopercle is devoid of spines in Sladenia and in other lophiiforms and paracanthopterygians. The remaining lophiid genera have a prominent spine at the base of the subopercle just anterior and slightly medial to the tip of the overlying opercle. Its presence is thus derived.
- Interopercular spines. All lophiids possess a dorsolaterally directed spine at the posterolateral angle of the interopercle. In Sladenia this spine is small; in the remaining genera it is enlarged and quite prominent. The presence of an enlarged, dorsolaterally directed interopercular spine is considered derived. In Lophiodes and Lophiomus there is a second posteriorly directed interopercular spine; this is occasionally double in some members of the Lophiodes mutilus species group. Since Lophiodes and Lophiomus share no other derived characters not also shared by Lophius, the presence of posterior interopercular spines must be considered a homoplasy.
- Gill opening. In all bony fishes except lophiiforms, the gill opening is anterior to the pectoral fin. In most lophiiforms, the gill opening is posterior to the pectoral fin and has been reduced to a rather narrow opening. The gill openings of lophiids are of two types: in Sladenia and Lophiodes the opening is very large and extends not only behind the pectoral fin, but in front of it as well; in Lophiomus and Lophius it is more restricted, located below and behind the pectoral fin. The state shared by Sladenia and Lophiodes is the primitive one and that shared by Lophiomus and Lophius is derived. This character, perhaps more than any other, supports Pietsch’s (1981) hypothesis that the lophiids are the primitive sister group of all other lophiiforms.
- Ural centrum and transverse process. In Sladenia, Lophiodes, other lophiiforms (except ogcocephalids), and other paracanthopterygians the ural centrum is round in cross section and devoid of transverse processes. In Lophiomus and Lophius, the ural centrum is strongly depressed and bears prominent transverse processes. The latter state is derived.
- Frontal ridges. In all lophiids and many other lophiiforms, each frontal bone bears a prominent ridge that runs forward from the eyes to the snout. This frontal ridge is smooth in Sladenia, Lophiodes, and all other lophiiforms that possess such a ridge. In Lophiomus and Lophius the frontal ridge is rugose, bearing short, sharp conical spines in Lophiomus and low, rounded knobs or ridges in Lophius. Th rugose ridge is thus derived.
- Sphenotic spines. All lophiids possess at least one dorsally directed spine on the sphenotic bone. In Sladenia and Lophiodes a single inner sphenotic spine is present. In Lophiomus and Lophius, an additional outer sphenotic spine is present (occasionally in Lophiomus a third posterior sphenotic spine is present). The presence of two sphenotic spines is thus derived.
- Epiotic spines. All lophiids possess a single spine on the epiotic bone. In Sladenia and Lophiodes the epiotic spine is small, while in Lophiomus and Lophius it is enlarged. The large epiotic spine is thus derived.
- Pectoral fin shape and number of pectoral-fin rays. In Sladenia and Lophiodes the pectoral fin is narrow and paddle-like, with a relatively low number of rays (18 or 19 in Sladenia, 13-22 in Lophiodes). In Lophiomus and Lophius, the pectoral fin is broad and fan-like, with a relatively high number of rays (21-25 in Lophiomus, 19-28 in Lophius). Outgroup comparison and analysis of the specialized function of the lophiid pectoral fin (Caruso, 1981:546) indicate the broad fin and high ray counts represent the derived condition.
- Number of vertebrae. Sladenia, Lophiodes, and Lophiomus have 18 or 19 vertebrae; Lophius has 26-31. The high vertebral counts are thus an autapomorphy of Lophius.
- Number of median-fin rays. Sladenia, Lophiodes, and Lophiomus have eight dorsal rays and six anal rays; Lophius has 9-12 dorsal rays and 8-10 anal rays. The high median-fin ray counts are thus an autapomorphy of Lophius.
- Upper quadrate spine. As stated above, Sladenia lacks quadrate spines, the remaining genera all have a single lower quadrate spine and Lophius has an additional upper quadrate spine. The presence of the latter is thus an autapomorphy of Lophius.
- Oral pigmentation. Most lophiid species have some diffuse pigment on the floor of the mouth between the oral valve and the lower pharyngeals. In Lophiomus there is a distinct reticular pattern of irregular anastomosing lines (in small specimens the pattern is more regular, appearing as pale circles on a dark background). This pigmentation is thus an autapomorphy of Lophiomus.
- Outer surface of maxilla and dentary. In Sladenia, Lophiodes, and Lophius, the external surfaces of the maxilla and dentary are smooth. In Lophiomus, short conical spines are present not only on the frontal ridge, but also on the external surfaces of the maxilla and dentary. The latter is thus considered an autapomorphy of Lophiomus.
- Posterior articular spines. As mentioned above, the presence of a single anterior articular spine is a synapomorphy of Lophiodes, Lophiomus, and Lophius. Lophiodes has an additional articular spine posterior to the jaw joint. The presence of this posterior articular spine is thus an autapomophy of Lophiodes.
- Third cephalic dorsal-fin spine. The third cephalic dorsal-fin spine is present in Lophiodes, Lophiomus, and Lophius, but absent in Sladenia. The absence of this spine in Sladenia is considered a loss and is thus an autapomorphy of Sladenia.
Distribution
Monkfish species are found in temperate, tropical, and subtropical waters of the Atlantic, Indian, and Western Pacific oceans. Monkfishes are peculiarly absent in the eastern Pacific. The genus Lophius is distributed on the continental shelf and slope of the Atlantic and off the coast of South Africa. The one species of Lophiomus is restricted to the Indian and Western Pacific oceans. Species of Lophiodes are distributed in the tropical and subtropical Atlantic, Indian, and Western Pacific. Members of the genus Sladenia are known from scattered localites in the western central Atlantic and Pacific.
Key to the Known Genera of the Family Lophiidae
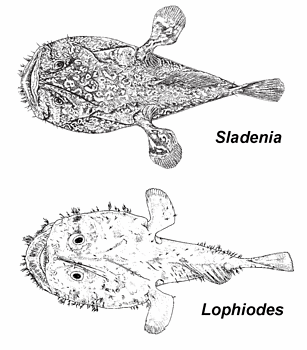
© 2005 John Caruso
- 1A. Gill opening extending below, behind, and in front of pectoral-fin base; frontal ridge smooth; inner sphenotic spine well developed, outer sphenotic spine represented by a low rounded knob; epiotic spine reduced (Go to 2)
- 1B. Gill opening extending below and behind, but not in front of pectoral-fin base; frontal ridge rugose; inner and outer sphenotic spines well developed; epiotic spine prominent (Go to 3)
- 2A. Head and body rounded, not depressed; third cephalic dorsal-fin spine absent; humeral, articular, quadrate, and subopercular spines absent (Sladenia Regan, 1908, Western Atlantic and Western Pacific oceans)
- 2B. Head and body depressed; third cephalic dorsal-fin spine present; humeral, articular, quadrate, and subopercular spines well developed (Lophiodes Goode and Bean, 1896, Atlantic, Pacific, and Indian oceans)
- 3A. Dorsal rays 8; anal rays 6; floor of mouth with dark reticulations; parietal spine reduced; a single quadrate spine; two interopercular spines; vertebrae 19 (Lophiomus Gill, 1882, Indo-West Pacific Ocean)
- 3B. Dorsal rays 9-12; anal rays 8-10; floor of mouth without dark reticulations; parietal spine prominent; two quadrate spines; a single interopercular spine; vertebrae 26-31 (Lophius Linnaeus, 1758, Atlantic, Western Pacific, and Indian oceans)
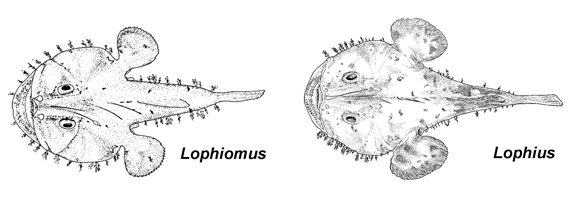
© 2005 John Caruso
References
Caruso, J. H. 1981. The systematics and distribution of the lophiid anglerfishes: I. A revision of the genus Lophiodes, with the description of two new species. Copeia, 1981(3): 522-549.
Caruso, J. H. 1983. The systematics and distribution of the lophiid anglerfishes: II. Revisions of the genera Lophiomus and Lophius. Copeia, 1983(1): 11-30.
Caruso, J. H. 1985. The systematics and distribution of the lophiid anglerfishes: III. Intergeneric relationships. Copeia, 1985(4): 870-875.
Pietsch, T. W. 1981. The osteology and relationships of the anglerfish genus Tetrabrachium, with comments on lophiiform classification. U. S. Fish. Bull., 79(3): 387-419.
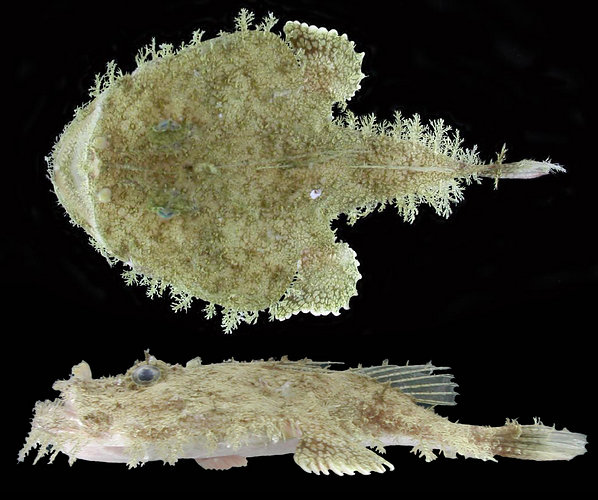
Title Illustrations

| Scientific Name | Lophiodes reticulatus Caruso & Suttkus |
|---|---|
| Location | Gulf of Mexico |
| Specimen Condition | Live Specimen |
| Identified By | J. H. Caruso |
| Life Cycle Stage | Adult |
| View | Dorsal and lateral |
| Collection | UF |
| Collector | USGS R/V Tommy Munro |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2002

|
About This Page
Tulane University, New Orleans, Louisiana, USA
Correspondence regarding this page should be directed to John H. Caruso at , Theodore W. Pietsch at , and Christopher P. Kenaley at
Page copyright © 2005
 Page: Tree of Life
Lophiidae. Goosefishes, monkfishes.
Authored by
John H. Caruso.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Lophiidae. Goosefishes, monkfishes.
Authored by
John H. Caruso.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 18 October 2005
Citing this page:
Caruso, John H. 2005. Lophiidae. Goosefishes, monkfishes. Version 18 October 2005 (under construction). http://tolweb.org/Lophiidae/21990/2005.10.18 in The Tree of Life Web Project, http://tolweb.org/

