



Marattiaceae
Maarten Christenhusz



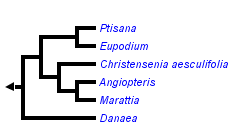


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Marattiaceae are represented by about 260 extant species and many fossil representatives. The extant members are placed in six genera: Angiopteris, Christensenia, Danaea, Eupodium, Marattia and Ptisana.
This early fern family is one of the oldest extant lineages of vascular plants that have changed little over periods of millions of years. They have a relatively good fossil record from the early Carboniferous (300 Ma) to the present day.
They can be easily distinguished from other ferns by their complex polycyclic stelar structure, often large, starchy rhizomes with fleshy or papery stipule-like outgrowths on each side of the petioles, presence of swollen nodes on rachises and often also on petioles, and eusporangia that are often fused and ligneous. All genera, except Christensenia, have free venation.
Marattiaceae are found in tropical or subtropical forests where the temperature and air humidity remain high throughout the year. While Angiopteris and Christensenia prefer lowland rain forest and grow in partly open habitats, Marattia has a preference for high elevation cloud forest and semi-deciduous forest in the subtropics. Danaea grows mostly in closed canopy, lowland and mountain rain forest and other shady, humid places, along stream banks, in ravines, and in sinkholes or near waterfalls, quite often on rather steep slopes.
Characteristics
The Marattiaceae are a distinct group of (almost always) terrestrial ferns with large, fleshy, erect or creeping rhizomes, with a polycyclic dictyostele (Sporne 1962, Smith et al. 2006), and mucilage canals in the roots, rhizomes and leaves. Young plants usually contain a mycorrhizal fungus - the oomycete Stigeosporium marattiacearum C. West - within the cortex (Sporne 1962). Other notable features are the starchy, leathery or papery stipule-like paired auricles at the base of each petiole, which are unique among ferns. Many species produce proliferous buds on the stipules or on the leaves (Uffelen 1994). The leaves are generally large and leathery (except in some “filmy” Danaea), and 1-3-pinnate (simple in some Danaea, or palmate in Christensenia). The petioles and rachises have polycyclic xylem and swollen pulvinae at the base of each internode. The sporangia are usually fused in round or elongate synangia, where a thin part of the sporangium dries and shrinks to form a pore through which the spores fall, except in Angiopteris where the sporangia are almost free. Each sporangium encloses very large numbers of spores (from about 1000 in Angiopteris to 7000 in Christensenia); these are usually monolete, bilateral or ellipsoid, and echinate (Sporne 1962, Smith et al. 2006).
The spores germinate rapidly, within a few days of being shed, and develop into large, monoecious, dark-green, mycorrhizal prothalli that can live for several years. An old prothallus may be several centimetres long and may closely resemble a large thalloid liverwort (Sporne 1962).
The young parts of Angiopteris, Ptisana, Eupodium and Marattia are covered with short simple hairs, while those of Christensenia and Danaea bear peltate scales. Microscopic characters, such as hairs, scales and indusia, which are of great taxonomic importance in other fern groups, are absent or vary little within marattioid genera.
Vegetative reproduction appears to be an important way of dispersal. Reproduction by means of stipule cuttings is also the easiest way of propagation in cultivation for Marattia and Angiopteris (Uffelen 1994, Chiou et al. 2006), but Christensenia and Danaea are difficult to cultivate because their requirements are unknown (Schneider 1893). Some Danaea species reproduce vegetatively, by means of apical buds on the leaves, where the terminal pinna is replaced or aborted; the bud can grow into a new plant when the leaf arches and the tip reaches the substrate. In this way, large uniform populations are sometimes formed. (After Christenhusz 2007)
Discussion of Phylogenetic Relationships
The first attempt to investigate evolutionary trends in Marattiales was published by Stidd (1974), who compared stelar structures among various fossil and extant genera. Hill and Camus (1986) examined generic relationships using a cladistic analysis of morphological characters, and proposed a new classification for Marattiales. In that study, they hypothesized that Christensenia was sister to a clade comprising Danaea, Marattia, and Angiopteris. Later phylogenetic studies based on DNA sequence data, which included some representatives from the Marattiaceae, consistently showed Danaea as sister to a clade uniting Marattia and Angiopteris (Hasebe et al. 1995, Pryer et al. 2004).
More recently molecular studies focussing on Danaea (Christenhusz et al. 2008) and Ptisana (Murdock 2008) have given greater insights into the indivisual relationships of this family.
The tree presented here is a consensus of these previous studies, which is inevitable because no good outgroup is available; all genera are too distant from the most closely related extant group, the Equisetaceae. Moreover not all species could be included in the sampling of the previous studies, and they are therefore here included based on their closest morphological resemblance.
References
Camus, J. M. (1990). Marattiaceae. In: Kubitzki, K, Kramer, K. U. and Green, P. S. (eds.). The Families and Genera of Vascular Plants, Vol. 1: Pteridophytes and Gymnosperms, pp. 174-180. Springer-Verlag, Berlin, New York.
Chiou, W. L., Huang, Y. M., and Chen, C. M. (2006). Conservation of two endangered ferns, Archangiopteris somai and A. itoi (Marattiaceae: Pteridophyta), by propagation from stipules. Fern Gaz. 17, pp. 271-278.
Christenhusz, M. J. M. (2007). Evolutionary History and Taxnomy of Neotropical Marattioid Ferns: Studies of an ancient lineage of plants. Annales Universitatis Turkuensis ser. AII, tom. 216, pp. 1-134.
Christenhusz, M. J. M., Tuomisto, H., Metzgar, J. S. and Pryer, K. M. (2008). Evolutionary relationships within the neotropical, eusporangiate fern genus Danaea (Marattiaceae). Molec. Phylog. Evol. 46, pp. 34-48.
Hasebe, M., Wolf, P. G., Pryer, K. M., Ueda, K., Ito, M., Sano, R., Gastony, G. J., Yokoyama, J., Manhart, J. R., Murakami, N., Crane, E. H., Haufler, C. H., and Hauk, W. D. (1995). Fern phylogeny based on rbcL nucleotide sequences. Amer. Fern J. 85, pp. 134-181.
Hill, C. R. and Camus, J. M. (1986). Evolutionary cladistics of marattialean ferns. Bull. Brit. Mus. (Nat. Hist.), Bot. 14, pp. 219-300.
Murdock, A. (2008a). Phylogeny of marattioid ferns (Marattiaceae): inferring a root in the absence of a closely related outgroup. Amer. J. Bot. 95, pp. 626-641.
Murdock, A. (2008b). A taxonomic revision of the eusporangiate fern family Marattiaceae, with description of a new genus Ptisana. Taxon 57, pp. 737-755.
Murdock, A. G., Reveal, J. L., and Doweld, A. (2006). Proposal to conserve Marattiaceae against Danaeaceae (Pteridophyta). Taxon 55: 1040-1042.
Presl, C. B. (1845). Marattiaceae. In: Genera Filicacearum, Supplementum Tentaminis Pteridographiae, pp. 7-40. Prague.
Pryer, K. M., Schuettpelz, E., Wolf. P. G., Schneider, H., Smith, A. R., Cranfill, R. (2004). Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences. Amer. J. Bot. 91, pp. 1582-1598.
Schneider, G. 1893. The Book of Choice Ferns for the Garden, Conservatory, and Stove, Vol. II from Blechnum to Nothchlaena. L. Upcott Gill, London, U.K.
Smith, A. R., Pryer, K. M., Schuettpelz, E., Korall, P., Schneider, H., and Wolf, P. G. (2006). A classification for extant ferns. Taxon 55, pp. 705-731.
Sporne, K. R. (1962). The morphology of pteridophytes, the structure of ferns and allied plants, pp. 127-135. Hutchison & Co. London.
Stidd, B. M. (1974). Evolutionary trends in Marattiales. Ann. Missouri Bot. Gard. 61, pp. 388-407.
Uffelen, G. van (1994). Varens, varens, varens, van Addertong tot Zwartsteel. Hortus Botanicus, Leiden, The Netherlands.
Underwood, L. M. (1909). Marattiaceae. North American Flora 16: 17-23.
Vriese, W. H. de, and Harting, P. (1853). Monographie des Marattiacées. Arnz, Leiden & Dusseldorf.
Information on the Internet
- Botany Photos: Marattiaceae. Plant Images by Maarten Christenhusz.
Title Illustrations

| Scientific Name | Angiopteris henryi |
|---|---|
| Location | Taiwan |
| Specimen Condition | Live Specimen |
| Source | 觀音座蓮舅科 Marattiaceae 台灣原始觀音座蓮 Archangiopteris henryi christ & Gies var. somai (Hay.) Tagawa. 96蓮華池 |
| Source Collection | Flickr |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike License - Version 2.0. This media file is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike License - Version 2.0.
|
| Copyright | © 2007 LiChieh Pan |

| Scientific Name | Ptisana salicina |
|---|---|
| Location | Auckland, New Zealand |
| Comments | Marattia salicina, King fern, Auckland, New Zealand. A large fern native to New Zealand and the South Pacific |
| Creator | Kahuroa |
| Specimen Condition | Live Specimen |
| Source | File:KingFernFrond.jpg |
| Source Collection | Wikimedia Commons |

| Scientific Name | Danaea mazeana Underw. |
|---|---|
| Location | Bains Jaunes, Basse-Terre, Guadeloupe |
| Specimen Condition | Live Specimen |
| Identified By | Maarten Christenhusz |
| Life Cycle Stage | sporophyte |
| Copyright |
© 2005 Maarten Christenhusz

|
About This Page
Maarten Christenhusz
The Natural History Museum, London, United Kingdom
Correspondence regarding this page should be directed to Maarten Christenhusz at
Page copyright © 2009 Maarten Christenhusz
 Page: Tree of Life
Marattiaceae.
Authored by
Maarten Christenhusz.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Marattiaceae.
Authored by
Maarten Christenhusz.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 23 January 2009
- Content changed 23 January 2009
Citing this page:
Christenhusz, Maarten. 2009. Marattiaceae. Version 23 January 2009. http://tolweb.org/Marattiaceae/134249/2009.01.23 in The Tree of Life Web Project, http://tolweb.org/


{kind=link}