



Myobatrachinae
Myobatrachine Frogs
David Cannatella

- Arenophryne
- Assa
- Crinia
- Geocrinia
- Indobatrachus

- Myobatrachus
- Paracrinia
- Pseudophryne
- Rheobatrachus
- Taudactylus
- Uperoleia
Introduction
This clade is endemic to Australia, New Guinea and Tasmania. Myobatrachines are generally smaller frogs with reduced ossification (See Ford and Cannatella, 1993). Myobatrachus, a head-first burrower and termite-eater that resembles Rhinophrynus, reaches 60 mm, but most of the other myobatrachines are 20-40 mm in snout-vent length. The gastric-brooding frog Rheobatrachus, which broods the developing eggs in its stomach, is also larger, 50-80 mm. It has been variously assigned to the Myobatrachinae, as its own subfamily or its own family.
The majority of myobatrachines are terrestrial or enjoy wet areas. Rheobatrachus is highly aquatic. Arenophryne is a burrowing frog in coastal sand dunes. Like its relative Myobatrachus, it burrows head-first. Some genera such as Crinia and Geocrinia inhabit swamps and wet areas. Taudactylus frequent mountain streams. Males of Assa have inguinal pouches in which larvae are reared.
The Eocene fossil †Indobatrachus has been included in this group (Spinar and Hodrová, 1985); a few Pleistocene fossils are known.
Geographic Distribution
The distribution of living members of the family Myobatrachinae is indicated in red.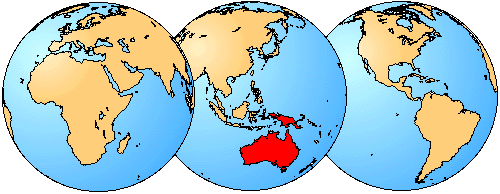
Discussion of Phylogenetic Relationships
Ford and Cannatella (1993) defined the node-based name Myobatrachinae to be the most recent common ancestor of Arenophryne, Assa, Crinia, Geocrinia, Myobatrachus, Paracrinia, Pseudophryne, Taudactylus, and Uperoleia, and all its descendants. Because the name is defined in terms of the ancestor and its descendants, it will remain stable regardless of whether Rheobatrachus or Sooglossidae is found to be nested within Myobatrachinae or not. Duellman and Trueb (1986) included the Eocene fossil †Indobatrachus in the Myobatrachinae; Ford and Cannatella (1993) did not use it in the definition because its relationships are uncertain. Rheobatrachus was not included in the definition, because it has been placed as closely related to Myobatrachinae, closely related to Limnodynastinae, or the sister-group of Limnodynastinae + Myobatrachinae.
Ford and Cannatella examined the literature on several character systems in myobatrachids (Heyer and Liem, 1976; Horton, 1982; Lynch, 1971) and found that in all instances myobatrachines and limnodynastines differed, and more importantly, that they shared no derived features that would unite them into a clade. Maxson (1992) summarized MC'F studies on myobatrachid frogs and concluded that there were two lineages corresponding to myobatrachine and limnodynastines, as had been adduced from morphological data. However, she did not compare these to other families, and thus could not make statements regarding the monophyly of Myobatrachidae.
Duellman and Trueb (1986) listed a ventral gap in the cricoid cartilage as uniting Myobatrachidae and Sooglossidae. However the ventral gap is present only in Myobatrachinae, not in Limnodynastinae (Lynch, 1973; Heyer and Liem, 1976). Lynch (1973) used that character to unite the (subfamily) Myobatrachinae and Sooglossidae, rendering Myobatrachidae paraphyletic. Heyer and Liem (1976) listed one unique feature uniting myobatrachines and limnodynastines to the exclusion of Rheobatrachus: the absence of prezygapophyses and transverse processes on the coccyx. However, coccygeal prezygapophyses and transverse processes are absent in leptodactylids (Lynch, 1971) and other neobatrachians, indicating that this character is incorrectly polarized.
Heyer and Liem (1976) reported two unique synapomorphies (among myobatrachids) for Myobatrachinae: the presence of notochordal vertebrae with free intervertebral discs and an incomplete cricoid ring. These were not present in Rheobatrachus or any limnodynastine (but see below under Sooglossidae). They listed three unique synapomorphies for Limnodynastinae: a connection between the intermandibularis and submentalis muscles, stalked alary processes (as opposed to broad and wing-like), anterior petrohyoid muscle inserting on the lateral edge of the hyoid (as opposed to insertion on the ventral surface of the hyoid).
Ford and Cannatella (1993) reasoned that the polarity of these latter two is reversed. Lynch (1971) reported that among leptodactylids and Heleophryne, the only taxa that share with Myobatrachinae the broad alary processes and ventral insertion of the anterior petrohyoid muscle are the leptodactylines Adenomera, Physalaemus, and Pseudopaludicola. Thus, the broad alary processes and ventral insertion of the anterior petrohyoideus are apomorphic within Neobatrachia. According to Heyer and Liem's (1976) data matrix, Rheobatrachus has a combination of the lateral insertion of the anterior petrohyoid (plesiomorphic) and a broad, wing-like alary process (derived). Assuming these data are correct, the latter of these two would unite Rheobatrachus with Myobatrachinae. Rheobatrachus is procoelous and has an entire cricoid ring; both of these are plesiomorphic. Another probable synapomorphy of Myobatrachinae is the reduction of the vomers and concomitant absence of vomerine teeth (Lynch, 1971).
Laurent (1979, 1986) recognized Rheobatrachus as a family distinct from the Myobatrachidae, based on its derived mode of reproduction--gastric brooding; most other workers (e.g., Heyer and Liem, 1976) have placed it in its own subfamily. Daugherty and Maxson (1982) found one-way immunological distances from Ranidella (=Crinia) signifera to other myobatrachines to be as great as 160 (Assa darlingtoni); the distance to Rheobatrachus silus was 133. They argued that Rheobatrachus was not more divergent from Ranidella than were some other myobatrachines. Farris et al. (1982) reanalyzed a subset of Heyer and Liem's (1976) data and found a myobatrachine clade and a limnodynastine clade, with Rheobatrachus as the sister-group to the limnodynastines, rather than the myobatrachines. However, reversing the polarity of the characters (discussed above) places Rheobatrachus closer to myobatrachines.
References
Click here for general list of references
Title Illustrations

| Scientific Name | Pseudophryne coriacea |
|---|---|
| Location | Tamborine National Park, Queensland, Australia |
| Specimen Condition | Live Specimen |
| Source | Red Backed Broodfrog |
| Source Collection | Flickr |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs License - Version 2.0. This media file is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs License - Version 2.0.
|
| Copyright | © 2008 teejaybee |
About This Page
If you are interested in authoring or co-authoring the page for this taxon, or some part of it (even a species), contact David Cannatella.
David Cannatella

University of Texas, Austin, Texas, USA
Correspondence regarding this page should be directed to David Cannatella at
Page copyright © 1995 David Cannatella
 Page: Tree of Life
Myobatrachinae. Myobatrachine Frogs.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Myobatrachinae. Myobatrachine Frogs.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Citing this page:
Cannatella, David. 1995. Myobatrachinae. Myobatrachine Frogs. Version 01 January 1995 (under construction). http://tolweb.org/Myobatrachinae/16946/1995.01.01 in The Tree of Life Web Project, http://tolweb.org/


