



Pteridaceae
Brake Ferns, Maidenhair Ferns, and allies
Carl Rothfels




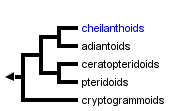


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Pteridaceae are one of the largest fern families, comprising over 1000 species (~10% of extant ferns). But even more notable than their species richness is the remarkable morphological and ecological diversity of the family, which includes obligate epiphytes, free-floating aquatics, brackish-swamp dwellers, several clades specialized for colonizing rocky deserts, and many generalist understory species. While some of these life forms do appear in other fern lineages (many other families, for example, contain epiphytes), most are rare and no other fern family contains such a wide range of ecological specializations.
Pteridaceae have been the subject of much taxonomic disagreement—their members have been distributed among at least 20 different families: Acrostichaceae; Actiniopteridaceae; Adiantaceae; Anopteraceae; Antrophyaceae; Bommeriaceae; Ceratopteridaceae; Cheilanthaceae; Coniogrammaceae; Crytopgrammaceae; Hemionitidaceae; Llaveaceae; Negripteridaceae; Notholaenaceae; Parkeriaceae; Platyzomataceae; Pteridaceae; Sinopteridaceae; Taenitidaceae; and Vittariaceae (Smith et al. 1996; Hassler and Swale, 2003). Some of these former family designations (e.g., Llaveaceae, Parkeriaceae, and Vittariaceae) correspond to monophyletic groups now known to be nested within the broader Pteridaceae, while others (e.g., Taenitidaceae) are assemblages of more distantly related species (Schuettpelz et al., 2007).
Characteristics
Perhaps due to the group's ecological breadth, Pteridaceae lack clear, unique, morphological features that unite their constituent species. One of the best characteristics is that members of this family never have true indusia—outgrowths of the lower leaf surface that protect the sporangia—although they frequently have modified leaf edges that serve the same function. These recurved leaf margins are sometimes strongly differentiated from the rest of the leaf tissue, and are thus referred to as “false indusia.”



Left: Abaxial leaf surfaces of Adiantum poiretii, showing sori and false indusia. © 2008 Carl Rothfels. Center: Fertile sporophyte of Coniogramme sp., showing sporangia distributed along the veins on the underside of the leaf. © 2007 . Right: Naked sporangia arranged along the veins of the leaf of Hemionitis palmata. © 2004
Instead of the round or elongate sori (clusters of sporangia) that are typical of many ferns, members of the Pteridaceae usually have their sporangia distributed along the veins on the underside of the leaf, often in marginal coenosori (a more-or-less contiguous line of sporangia around the margin of the leaf). More rarely, the sporangia may be attached to the false indusium itself (Adiantum), or spread evenly across the undersurface of the leaf (Acrostichum). In addition to these macromorphological characteristics, members of the Pteridaceae tend to share a chromosome base number of x=30 (occasionally x=27, 28, or 29).
Discussion of Phylogenetic Relationships
Plastid DNA sequence data suggest that the Pteridaceae comprise five (mostly) well-supported clades (Schuettpelz et al., 2007). The cheilanthoids are sister to the adiantoids (which include both Adiantum and the group formerly referred to as the Vittariaceae); these two clades are together sister to the pteridoids plus the ceratopteroids; and the cryptogrammoids are, in turn, sister to this large clade. Of these five clades, only the cheilanthoids lack strong support, due to some uncertainty about the phylogenetic position of their earliest-diverging branch (the Doryopteris ludens clade—see the cheilanthoid page). These five clades, none of which correspond perfectly with previously recognized families or subfamilies, do show some ecological differentiation. The ceratopteroids contain the aquatic taxa; the pteridoids contain most of the generalist forest understory species; and the cheilanthoids contain most of the xeric habitat specialists.
This phylogenetic structure differs from the most recent formal taxonomic treatments of the family (Kramer, 1990; Tryon et al., 1990), which recognized two families (Pteridaceae and Vittariaceae), and divided the Pteridaceae into six subfamilies: Adiantoideae, Ceratopteridoideae, Cheilanthoideae, Platyzomatoideae, Pteridoideae, and Taenitidoideae. Molecular data demonstrate that the Vittariaceae are nested within Pteridaceae (specifically within the adiantoid clade), and, while the circumscriptions may be close in some cases, none of the formally described subfamilies of Tryon and Tryon (1990; Adiantoideae, Cheilanthoideae, etc.) corresponds perfectly to the informal clades recognized here (adiantoids, cheilanthoids, etc.). A formal revision of Pteridaceae awaits further data.
"Pteroids" vs. "Pteridoids"
Note that the term “pteroid” is colloquial shorthand to refer to members of the Pteridaceae, whereas “pteridoid” refers to members of a specific Pteridaceae subclade.
References
Benham, D. M., and M. D. Windham. 1992. Generic affinities of the star-scaled cloak ferns. American Fern Journal 82:47-58.
Brown, R. 1810. Prodromus Florae Novae Holandiae et Insulae Van-Diemen. Richard Taylor & Sons, London.
Copeland, E. B. 1947. Genera Filicum. Chronica Botanica Co., Waltham.
Gastony, G. J., and D. R. Rollo. 1995. Phylogeny and generic circumscriptions of cheilanthoid ferns (Pteridaceae: Cheilanthoideae) inferred from rbcL nucleotide sequences. American Fern Journal 85:341-360.
Gastony, G. J., and D. R. Rollo. 1998. Cheilanthoid ferns (Pteridaceae: Cheilanthoideae) in the southwestern United States and adjacent Mexico--A molecular phylogenetic reassessment of generic lines. Aliso 17:131-144.
Giauque, M. F. A. 1949. Wax glands and prothallia. American Fern Journal 39:33-35.
Hall, C. C. 1950. Notholaena copelandii, a newly recognized species of the Texano-Mexican region. American Fern Journal 40:178--187.
Hassler, M., and B. Swale. 2003. Checklist of World Ferns. http://homepages.caverock.net.nz/~bj/fern/
Hevly, R. H. 1963. Adaptations of cheilanthoid ferns to desert environments. Journal of the Arizona Academy of Science 2:164-175.
Kirkpatrick, R. E. B. 2007. Investigating the monophyly of Pellaea (Pteridaceae) in the context of a phylogenetic analysis of cheilanthoid ferns. Systematic Botany 32:504-518.
Kramer, K. U. 1990. Notes on the higher level classification of the recent ferns. Pages 49--52 in The Families and Genera of Vascular Plants (K. U. Kramer, and P. S. Green, eds.). Springer-Verlag, Berlin.
Lellinger, D. B. 1989. The ferns and fern-allies of Costa Rica, Panama, and the Chocó (Part 1: Psilotaceae through Dicksoniaceae). Pteridologia 2A:1-364.
Mickel, J. T. 1979. The fern genus Cheilanthes in continental United States. Phytologia 41:431-437.
Mickel, J. T., and A. R. Smith. 2004. The Pteridophytes of Mexico. The New York Botanical Garden Press, New York.
NatureServe. 2008. NatureServe Explorer, Arlington, Virginia. http://www.natureserve.org/explorer/
Pichi Sermolli, R. E. G. 1989. Again on the typification of the generic name Notholaena R. Brown. Webbia 43:301-310.
Prado, J., C. D. N. Rodrigues, A. Salatino, and M. L. F. Salatino. 2007. Phylogenetic relationships among Pteridaceae, including Brazilian species, inferred from rbcL sequences. Taxon 56:355-368.
Rothfels, C. J., M. D. Windham, A. L. Grusz, G. J. Gastony, and K. M. Pryer. 2008. Toward a monophyletic Notholaena (Pteridaceae): Resolving patterns of evolutionary convergence in xeric-adapted ferns Taxon 57:712-724.
Schuettpelz, E., H. Schneider, L. Huiet, M. D. Windham, and K. M. Pryer. 2007. A molecular phylogeny of the fern family Pteridaceae: assessing overall relationships and the affinities of previously unsampled genera. Molecular Phylogenetics and Evolution 44:1172-1185.
Seigler, D. S., and E. Wollenweber. 1983. Chemical variation in Notholaena standleyi. American Journal of Botany 70:790-798.
Tryon, R. M. 1956. A revision of the American species of Notholaena. Contributions from the Gray Herbarium 179:1-106.
Tryon, R. M., and A. F. Tryon. 1973. Geography, spores, and evolutionary relations in the cheilanthoid ferns. Pages 145-153 in The phylogeny and classification of ferns. (A. C. Jermy, J. A. Crabbe, and B. A. Thomas, eds.). Academic Press, New York.
Tryon, R. M., and A. F. Tryon. 1982. Ferns and Allied Plants with Special Reference to Tropical America. Springer-Verlag, New York.
Tryon, R. M., A. F. Tryon, and K. U. Kramer. 1990. Pteridaceae. Pages 404 in The Families and Genera of Vascular Plants (K. U. Kramer, and P. S. Green, eds.). Springer-Verlag, Berlin.
Windham, M. D. 1987. Argyrochosma, a new genus of cheilanthoid ferns. American Fern Journal 77:37-41.
Windham, M. D. 1993a. Notholaena. Pages 143--149 in Flora of North America (Flora of North American Editorial Committee, ed.) Oxford University Press, New York.
Windham, M. D. 1993b. Pteridaceae. Pages 122--124 in Flora of North America (Flora of North American Editorial Committee, ed.) Oxford University Press, New York.
Windham, M. D., L. Huiet, E. Schuettpelz, C. J. Rothfels, J. Beck, A. L. Grusz, G. Yatskievych, and K. M. Pryer. 2008. Using plastid and nuclear DNA sequences to redraw generic boundaries and demystify species complexes in cheilanthoid ferns. American Fern Journal (in review).
Windham, M. D., and G. Yatskievych. 2003. Chromosome studies of cheilanthoid ferns (Pteridaceae: Cheilanthoideae) from the western United States and Mexico. American Journal of Botany 90:1788-1800.
Wollenweber, E. 1984. Exudate flavonoids of Mexican ferns as chemotaxonomic markers. Rev. Latinoamer. Quim. 15:3-11.
Wollenweber, E., and H. Schneider. 2000. Lipophilic exudates of Pteridaceae -- chemistry and chemotaxonomy. Biochemical Systematics and Ecology 28:751-777.
Yatskievych, G., and A. L. A. Arbeláez. 2008. A new species and three generic transfers in the fern genus Notholaena (Pteridaceae). Novon 18:120-124.
Yatskievych, G., and A. R. Smith. 2003. Typification of Notholaena R. Br. (Pteridaceae). Taxon 52:331-336.
Yatskievych, G., M. D. Windham, and E. Wollenweber. 1990. A reconsideration of the genus Pityrogramma (Adiantaceae) in western North America. American Fern Journal 80:9.
Zhang, G., X. Zhang, and Z. Chen. 2005. Phylogeny of cryptogrammoid ferns and related taxa based on rbcL sequences. Nordic Journal of Botany 23:485-493.
Title Illustrations

| Scientific Name | Scoliosorus ensiformis |
|---|---|
| Location | Costa Rica, Cuerici Biological Station |
| Specimen Condition | Live Specimen |
| Identified By | R.Moran |
| Life Cycle Stage | Sporophyte, fertile |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 Carl Rothfels

|

| Scientific Name | Astrolepis integerrima |
|---|---|
| Location | Texas |
| Specimen Condition | Live Specimen |
| Identified By | Michael Windham |
| Life Cycle Stage | Sporophyte, fertile |
| Collector | J.Beck |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 Carl Rothfels
|

| Scientific Name | Cryptogramma crispa |
|---|---|
| Location | Canada, British Columbia |
| Specimen Condition | Live Specimen |
| Identified By | C.J.Rothfels |
| Life Cycle Stage | sporophyte |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 Carl Rothfels
|

| Scientific Name | Acrostichum danaeifolium |
|---|---|
| Location | Costa Rica, Prov. de Guanacaste, Palo Verde National Park |
| Comments | A fine sterile leaf being modeled by Mauricio Bonifacino. |
| Acknowledgements | Photo reproduced with permission from PlantSystematics.org |
| Specimen Condition | Live Specimen |
| Identified By | R.Moran |
| Life Cycle Stage | Sporophyte, sterile leaf |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright | © 2006 |
About This Page
Carl Rothfels
Duke University, Durham, North Carolina, USA
Correspondence regarding this page should be directed to Carl Rothfels at
Page copyright © 2009 Carl Rothfels
 Page: Tree of Life
Pteridaceae . Brake Ferns, Maidenhair Ferns, and allies.
Authored by
Carl Rothfels.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Pteridaceae . Brake Ferns, Maidenhair Ferns, and allies.
Authored by
Carl Rothfels.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 23 December 2008
- Content changed 23 December 2008
Citing this page:
Rothfels, Carl. 2008. Pteridaceae . Brake Ferns, Maidenhair Ferns, and allies. Version 23 December 2008 (under construction). http://tolweb.org/Pteridaceae/29352/2008.12.23 in The Tree of Life Web Project, http://tolweb.org/

