




Somasteroidea
Alexander E. Primus



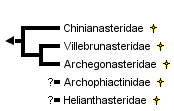


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxCladogram based on arguments from Spencer and Wright (1966), McKnight (1975), and Blake (1982).
Introduction
Somasteroidea are an extinct subclass of asterozoan echinoderms (star-shaped echinoderms), known to have lived during the early Paleozoic (Spencer, 1951). Somasteroids first appear in the fossil record in the Early Ordovician (Tremadocian, ~510-493 Ma), just prior to the appearance of the other two asterozoan classes, Asteroidea (sea stars) and Ophiuroidea (brittle stars). While the asteroids and ophiuroids have subsequently undergone extensive radiations and represent the two most successful extant echinoderm classes, somasteroid forms do not appear in the fossil record after the late Devonian and are therefore thought to have been extinct since then (Spencer, 1951; Blake, 1982).
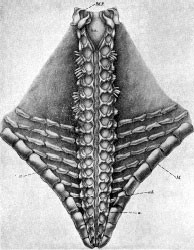
Somasteroids are flattened, fivefold radially symmetrical echinoderms that range in shape from that of a pentagon to that of a star with protruding arms. It appears that somasteroids were benthic marine animals that were filter- and/or detritus-feeders. Despite their relatively rigid arm structure, somasteroids are thought to have been somewhat mobile and able to scavenge along the ocean floor (Spencer, 1951).
Characteristics
Spencer's (1951) original organization of the early asterozoans was primarily based upon the activities of the arms with respect to feeding. While early asteroids had stout and muscular arms, adapted for a carnivorous diet of large food items, and early ophiuroids had long and narrow arms, adapted for feeding on small food particles present in or near the sea floor, the broad, undifferentiated, petaloid arms of somasteroids appeared to have had little or no role in feeding. More contemporary classification schemes of the asterozoans have relied upon the subsequent diversification of certain asterozoan arm morphological characteristics, such as the types and organization of arm skeletal elements. In the following description of somasteroid morphology, comparisons to asteroid and ophiuroid morphology will be made in an effort to signify some of the key defining characteristics.
Asterozoans, as described by Spencer (1951), have an appreciably flattened central body cavity from which five arms extend. As in asteroids, the arms and body of somasteroids are fully continuous. The arms and the disk-like body of ophiuroids, on the other hand, are clearly discontinuous. The two flat sides of asterozoans comprise the oral and aboral surfaces, of which the oral side is in contact with the substrate. A disjunct ossicular skeleton called a test covers both oral and aboral surfaces of asterozoans. Adjacent skeletal ossicles of the test are not rigidly fused as they are in crinoids and most echinoids, but are joined with connective and muscular tissues. The ossicular skeleton of somasteroids is often refered to as simple or undifferentiated when compared to that of asteroids and ophiuroids. Nevertheless, while the aboral test of somasteroids is composed of a single ossicle type (paxillae with tetraradiate bases), the oral test is significantly more complex (see Figures 1 and 2).
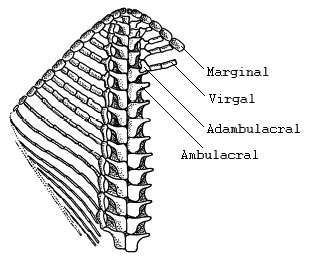
Figure 1. Interpretation of one arm of Ampullaster ubaghasi (Villebrunasteridae), view of oral test.
Figure modified from Fell (1963b). Drawing by H. B. Fell. Copyright © 1963 Royal Society of London.
The oral surface of each arm consists of two adjacent rows of recumbent ambulacral plates (ambulacrals). These ambulacrals, presumably involved in feeding, extend from the centrally located mouth to the tip of each arm. In asteroids, ophiuroids and most somasteroids, adjacent ambulacrals are arranged in a paired manner. In some early somasteroids, however, adjacent ambulacrals are alternately arranged. Each ambulacral gives rise to a transverse series of ossicles (metapinnules). These ossicles can occur as undifferentiated rod-like elements called virgals (found only in somasteroids), or as various differentiated elements including adambulacrals, intermediate ossicles, and marginal ossicles. When present in somasteroids and ophiuroids, adambulacrals are positioned laterally with respect to ambulacrals. In asteroids, however, adambulacrals are located underneath the ambulacrals. In addition, in asteroids the ambulacral plates are raised slightly off the surface upon which they sit, creating a space between the ambulacrals and the substrate. This space is often refered to as the ambulacral furrow or groove. Somasteroids and ophiuroids lack a permanent ambulacral furrow, although it appears that some derived somasteroids may have been able to create a temporary ambulacral furrow through muscular flexures.
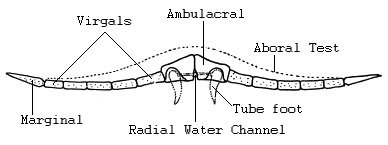
Figure 2. Reconstructed cross-section of one arm of Chininanaster sp. (Chinianasteridae).
Figure modified from Fell (1963b). Drawing by H. B. Fell. Copyright © 1963 Royal Society of London.
Radial water vessels extend the length of each arm within a radial channel formed by the recumbent ambulacral plates and give rise to numerous tube feet (Figure 2). In somasteroids, the radial water vessel is often positioned at the oral-marginal junction of ambulacrals, as in asteroids, but can also be enclosed within the ambulacral plates, as in ophiuroids. Tube feet branching from the radial water vessels are seated in broad basins that, in some species, also give rise to ampulae that extend into the body cavity through ambulacral pores. The jaw ossicles of somasteroids are differentiated with small mouth angle plates, in contrast to those of asteroids that lack odontophores (McKnight, 1975). Buccal slits, which function primarily as respiratory structures in ophiuroids, may or may not have been present adjacent to the mouth frame in somasteroids.
Origin of Somasteroidea
Discussions addressing somasteroid origins have certainly been a major point of contention through the years primarily because the intermediate forms that gave rise to the somasteroids are lacking from the fossil record (Blake and Guensburg 1993; Blake, 1994). It seems most likely, however, that the somasteroids emerged from either a crinoid lineage (Fell, 1963; Mooi and David, 2000), or an edrioasteroid lineage (Paul and Smith, 1984; Smith 1988b).
Discussion of Phylogenetic Relationships
Informative evolutionary and phylogenetic analyses within Somasteroidea have been quite difficult because the examination and characterization of the group has been somewhat limited. What is known about the diversification of somasteroids, however, is primarily derived from work on three lower Ordovician families: Chinianasteridae (Tremadoc-Early Arenig), Villebrunasteridae (Tremadoc-Early Arenig), and Archegonasteridae (Late Arenig), as discussed by Nichols (1962), Fell (1963), and Blake (1982).
The Family Chinianasteridae is thought to include the most basal group of somasteroids based on the presence of undifferentiated virgals and robust ambulacrals, in addition to the absence of ampulae. The more derived Family Villebrunasteridae has differentiated virgals that include adambulacral, oral intermediate, and distinct marginal elements. Members of this group also have internal ampulae that provide a more direct connection between the body cavity and the environment. The most derived of these three families, the Archegonasteridae, represent the most thoroughly examined group of somasteroids. The Archegonasteridae have undergone a virgal reduction and have changed their body shape slightly from their predecessors to a more or less pentagonal outline with no interradial slits. In addition, Archegonasteridae specimens appear to have a more robust mouth frame than the more basal somasteroids and are often preserved with an ambulacral furrow. The latter two of these Archeogasteridae characteristics are also quite typical of asteroids and may very well have facilitated an adaptive change in feeding behavior from scavenging to predation.
Classification of Somasteroidea
The first organized classification of Somasteroidea was carried out by W. K. Spencer (1951). In his report, Spencer describes some of the earliest asteroid and ophiuroid specimens ever found. In addition, he describes fossils of a third asterozoan group, the somasteroids, which occur not only contemporaneously with, but also prior to all recognizable asteroid and ophiuroid fossil forms. These findings are suggestive of a scenario in which all asterozoans arose from ancient somasteroid forms.
Spencer (1951) assigned one order, Goniactinida, to the subclass Somasteroidea. Within this order he assigned two families, Chinianasteridae, including the genera Chinianaster (Tremadoc-Early Arenig) and Villebrunaster (Tremadoc-Early Arenig), and Archegonasteridae, composed of one genus, Archegonaster (Late Arenig). Subsequently, a seemingly extant somasteroid was identified by Fell (1962) in the Pacific off the coast of southern Mexico. This extant specimen, Platasterias latiradiata, was placed into a new somasteroid family, Platasteriidae (Fell, 1963a). The following year, Fell (1963b) reclassified the entire Asterozoa, and in so doing, removed Villebrunaster from the family Chinianasteridae and placed it in its own family Villebrunasteridae. Spencer and Wright (1966) added one more family, Archophiactinidae composed of three genera, Archophiactis (Late Ordovician), Stuertzurea (Silurian), and Lepidasterina (Late Devonian). McKnight (1975) then placed one more family, Helianthasteridae, previously an asteroid group, into Somasteroidea. The extant Platasterias latiradiata was subsequently reclassified by D. B. Blake (1982) and was placed in the Asteroidea. Platasteriidae was consequently removed from the somasteroids. This left five families and between 8-12 genera in the subclass Somasteroidea (Smith and Jell, 1990), as can be seen in the cladogram, based on Spencer and Wright (1966), McKnight (1975), and Blake (1982), shown above.
References
Blake, D.B. 1982. Somasteroidea, Asteroidea, and the Affinities of Luidia (Plasterias) Latiradiata. Paleontology, 25:167-191.
Blake, D.B. 1994. Re-evaluation of the Palasteriscidae Gregory, 1900, and the early phylogeny of the Asteroidea (Echinodermata). J. Paleont. 68:123-134.
Blake, D.B. 1998. Morphological characters of early asteroids and ophiuroids. Echinoderms: San Francisco, Mooi and Telford (eds.), Balkema, Rotterdam.
Blake, D.B., Guensburg, T.E., 1993. New Lower and Middle Ordovician stelleroids (Echinodermata) and their bearing on the origins and early history of the stelleroid echinoderms.
Fell, H.B. 1962. A surviving somasteroid from the eastern Pacific Ocean. Science, N.Y. 136:633-636.
Fell, H.B. 1963a. A new family and genus of Somasteroidea. Trans. R. Soc. N. Z. 3:143-146.
Fell, H.B. 1963b. The phylogeny of sea-stars. Phil. Trans. R. Soc. Lond., ser. B 246:381-435.
McKnight, D.G. 1975. Classification of Somasteroids and Asteroids (Asterozoa: Echinodermata). J. Roy. Soc. N. Z. 5:13-19.
Mooi, R., David, B. 2000. What a new model of skeletal homologies tells us about asteroid evolution. Amer. Zool. 40:326-339.
Nichols, D. 1962. Echinoderms. Huchinson and Co., London.
Paul, C.R.C., Smith, A.B. 1984. The early radiation and phylogeny of echinoderms. Biological Reviews, 46:157-200.
Smith, A.B. 1988. Patterns of diversification and extinction in early Paleozoic echinoderms. Paleontology, 31:799-828.
Smith, A.B., Jell, P.A. 1990. Cambrian ederoasteroids from Australia and the origins of starfishes. Mem. Queen. Mus. 28:715-778.
Spencer, W.K. 1951. Early Paleozoic starfish. Phil. Trans. R. Soc. Lond., ser. B 235:87-129.
Spencer, W.K., Wright, C.W. 1966. Asterozoans, pp. S4-S107. In R.C. Moore (ed.), Treatise in Invertebrate Paleontology, Part U, Echinodermata 3. Geological Society of America and the University of Kansas Press, New York and Lawerance.
Title Illustrations

| Scientific Name | Villebrunaster thorali (Villebrunasteridae) |
|---|---|
| Location | from south France |
| Reference | After Spencer, W.K. 1951. Early Paleozoic starfish. Phil. Trans. R. Soc. Lond., ser. B 235:87-129. |
| Specimen Condition | Fossil -- Period: Lower Ordovician, Tremadoc-Early Arenig |
| Body Part | ventral side showing schematic of oral test |
| Copyright | © 1951 Royal Society of London |

| Scientific Name | Archegonaster pentagonus (Archegonasteridae) |
|---|---|
| Location | from former Czechoslovakia |
| Reference | After Spencer, W.K. 1951. Early Paleozoic starfish. Phil. Trans. R. Soc. Lond., ser. B 235:87-129. |
| Specimen Condition | Fossil -- Period: Lower Ordovician, Late Arenig |
| Body Part | ventral side showing schematic of oral test |
| Copyright | © 1951 Royal Society of London |
| Scientific Name | Chinianaster levyi (Chinianasteridae) |
|---|---|
| Location | from south France |
| Reference | After Spencer, W.K. 1951. Early Paleozoic starfish. Phil. Trans. R. Soc. Lond., ser. B 235:87-129. |
| Specimen Condition | Fossil -- Period: Lower Ordovician, Tremadoc-Early Arenig |
| Body Part | ventral side showing schematic of oral test |
| Copyright | © 1951 Royal Society of London |
About This Page

Duke University, Durham, North Carolina, USA
Correspondence regarding this page should be directed to Alexander E. Primus at
Page copyright © 2005
 Page: Tree of Life
Somasteroidea.
Authored by
Alexander E. Primus.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Somasteroidea.
Authored by
Alexander E. Primus.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 05 January 2005
Citing this page:
Primus, Alexander E. 2005. Somasteroidea. Version 05 January 2005. http://tolweb.org/Somasteroidea/24272/2005.01.05 in The Tree of Life Web Project, http://tolweb.org/

