



Vertebrata
Animals with backbones
Philippe Janvier
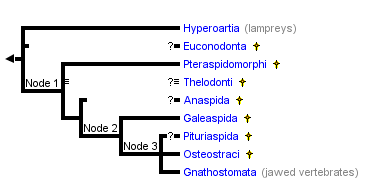


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxThe main characteristics supporting the nodes of this phylogeny are:
Node 1: Mineralized exoskeleton, sensory-line canals and grooves
Node 2: Perichondral bone or calcification, externally open endolymphatic duct
Node 3: Paired fins containing musculature and concentrated in pectoral position, two dorsal fins, epicercal (i.e. upwardly tappering) tail, sclerotic ring and scleral ossification, cellular dermal bone
Introduction
The Vertebrata, or vertebrates, is a very diverse group, ranging from lampreys to Man. It includes all craniates, except hagfishes, and are characterized chiefly by a vertebral column, hence their name. The majority of the extant vertebrates are the jawed vertebrates, or gnathostomes, but lampreys are jawless vertebrates. However, in Late Silurian or Early Devonian times, about 420 to 400 million years ago, the situation was reverse, and the majority of the vertebrate species were jawless fishes (the "ostracoderms", presumably more closely related to the gnathostomes than to lampreys). The decline of the jawless vertebrates and the subsequent rise of the gnathostomes took place about 380 million years ago.
Extant vertebrates comprise two clades: the Hyperoartia, or lampreys, and the Gnathostomata, or jawed vertebrates. In addition, there is a number of taxa of fossil jawless vertebrates which were formerly referred to as the "ostracoderms" ("shell-skinned") because most of them possess an extensive, bony endo- and exoskeleton. The "ostracoderms" lived from the Early ordovician (about 480 million years ago) to the Late Devonian (about 370 million years ago). The relationships of the various groups of "ostracoderms" has been the subject of considerable debate since the mid-nineteenth Century, and the theory of relationship proposed here is far from definitive, yet the best supported by the currently available data. The "ostracoderms" are represented by five major groups, four of which are almost certainly clades: the Heterostraci, Osteostraci, Galeaspida, Anaspida, and Thelodonti (the monophyly of the latter being debated, Thelodonti page). In addition, there are minor groups which only include a few species: the Arandaspida, Astraspida, Eriptychiida, and Pituriaspida. The Arandaspida, Astraspida, Eriptychiida, and Heterostraci are regarded as forming a clade, the Pteraspidomorphi. Some monospecific genera, Jamoytius, Endeiolepis, and Euphanerops, formerly referred to the Anaspida, are now removed from that clade and may be more closely related to lampreys (see Hyperoartia). A large but still poorly known group, the Euconodonta, has recently been included in the Craniata, and possibly the Vertebrata. It is currently referred to as 'conodonts', but the only forms that can reliably be regarded as craniates belong to a subgroup of conodonts known as euconodonts.
Characteristics
The Vertebrata have all the characteristics of the Craniata but share, in addition, a number of unique characteristics which do not occur in hagfishes (Hyperotreti). These characteristics are:
- Metamerically arranged endoskeletal elements flanking the spinal cord. There are primitively two pairs of such elements in each metamere and on each side: the interdorsals and basidorsals. In the gnathostomes, there are two additional pairs ventrally to the notochord: the interventrals and basiventrals. These elements are called arcualia and can fuse to a notochordal calcification, the centrum. The ensemble of the arcualia , centrum is the vertebra, and the ensemble of the vertebrae is the vertebral column.
Click on an image to view larger version & data in a new window
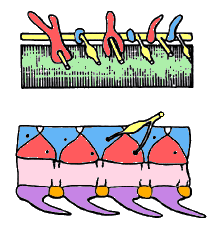
The vertebrates are characterized by a vertebral column; that is, a variable number of endoskeletal elements aligned along the notochord (green) and flanking the spinal cord (yellow). In lampreys (top), the vertebral elements are only the basidorsal (red) and the interdorsals (blue). In the gnathostomes, there are in addition ventral elements, the basiventrals (purple) and interventrals (orange), and the notochord may calcify into centra (pink). (After Janvier 1996).
- Extrinsic eye muscles. These muscles are attached to the eyeball and orbital wall, and ensure eye movements
- Radial muscles in fins. These are small muscles associated with each of the cartilaginous radials of the unpaired and paired fins. They ensure the undulatory movements of the fin web.
- Atrium and ventricle of heart closely-set.
- Nervous regulation of heart. The heart in the embryo of the vertebrates is aneural, like the heart of adult hagfishes. In adult vertebrates, however, the heart is innervated by a branch of the vagus nerve.
- Typhlosole in the intestine. This is a spirally coiled fold of the intestinal wall. In the Gnathostomes, it can be developed into a complex spiral valve.
- At least two vertical semicircular canals in the labyrinth
- True neuromasts in the sensory-line system
There are many other vertebrate characteristics, both anatomical and physiological.
Discussion of Phylogenetic Relationships
As for extant vertebrates, the main question is whether lampreys are the sister-group of the gnathostomes, or that of hagfishes. In the latter case there would be no reason to distinguish the Vertebrata from the Craniata, as it was formerly done. Although there is good evidence for the lamprey-gnathostome sister-group relationship, the theory that the cyclostomes (lampreys,hagfishes) are a clade is still supported by a number of zoologists. Considering the large number of anatomical, physiological and molecular data that are available now to test these theories, one can expect a definitive clue in a near future (for discussion, see Craniata).
The question of the relationships of the numerous extinct vertebrate groups is, in contrast, far from being resolved. This chiefly concerns the Palaeozoic taxa formerly referred to as "ostracoderms"; that is, armored jawless craniates, which are likely to be vertebrates and are now considered as being all more closely related to the gnathostomes than to lampreys.
During most of the nineteenth century, the "ostracoderms" known at that time (i.e. the Heterostraci and Osteostraci) were regarded as bony fishes, until Cope (1889) suggested to include them with lampreys and hagfishes in the taxon Agnatha ("jawless"). In the beginning of the twentieth century, Kiaer (1924) and Stensiö (1927) showed that the Anaspida and Osteostraci share with lampreys a median, dorsally placed "nostril" (in fact a nasohypophysial opening) and suggested to include these three groups in a clade Cephalaspidomorphi. In addition, Stensiö (1927) proposed that hagfishes were derived from the Heterostraci and should be grouped with them in the Pteraspidomorphi. At that time, however, the Agnatha were regarded as a clade, whose sister-group was the Gnathostomata, as illustrated by Stensiö's (1927) diagram:
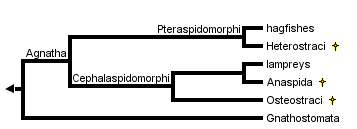
This theory implied the diphyletic origin of the Recent "cyclostomes" (hagfishes and lampreys). Although they accepted the monophyly of the Cephalaspidomorphi, most paleontologists rejected that of the Pteraspidomorphi (as including hagfishes). In contrast, until the 1970's, it was widely accepted that the Heterostraci are more closely related, or ancestral to the gnathostomes, mainly because they lacked the specializations of the Cephalaspidomorphi and because they had paired olfactory capsules, like the gnathostomes. With the rise of cladistics, in the late 1970's and the 1980's, and following Løvtrup's (1977) suggestion that extant cyclostomes were paraphyletic, a number of trees were published, which all showed the "ostracoderms" (and the Agnatha as a whole) as paraphyletic. However, all these trees implied that lampreys had lost several characteristics, in particular the paired fins, mineralized skeleton, and sensory-line canals. A major change was Gagnier's (1993) first computer-generated tree, in which these reversions were avoided by considering all "ostracoderms" as more closely related to the gnathostomes than to either lampreys and hagfishes. Further analyses (Forey & Janvier 1994, Janvier 1996b) largely confirmed the higher degree of parsimony of this phylogeny. Although there are variations as to the position of certain taxa, the Galeaspida and Osteostraci constantly group together with the Gnathostomes, whereas the Astraspida, Eriptychiida, Arandaspida, and Heterostraci form a clade, the Pteraspidomorphi, albeit poorly supported. One of the consequences of this tree is that the dorsal nasohypophysial opening (formerly the characteristic of the Cephalaspidomorphi) either occurred more than once, or is a general feature of the Vertebrata.
In this tree, four fossil groups are positioned with a question mark. In the case of the Euconodonta, Anaspida and Pituriaspida, this uncertainty is largely due to the scarcity of the characters available from the material (in particular as to the internal anatomy). In the case of the Thelodonti, it is due to their controversial status, as they are likely to be a paraphyletic assemblage of stem Heterostraci and possibly stem forms of other "ostracoderm" groups, yet some authors regard them as a clade (see Thelodonti page).
References
Forey, P. L. (1984). Yet more reflections on agnathan-gnathostome relationships. Journal of Vertebrate Paleontology, 4, 330-343.
Forey, P. L., and Janvier, P. (1993). Agnathans and the origin of jawed vertebrates. Nature, 361, 129-134.
Forey, P. L., and Janvier, P. (1994). Evolution of the early vertebrates. American Scientist, 82, 554-565.
Hardisty, M. W. (1982). Lampreys and hagfishes: Analysis of cyclostome relationships. In The Biology of Lampreys, (ed. M. W. Hardisty and I. C. Potter), Vol.4B, pp. 165-259. Academic Press, London.
Janvier, P. (1993). Patterns of diversity in the skull of jawless fishes. In The skull (ed. J. Hanken and B. K. Hall), Vol. 2, pp. 131-188. The University of Chicago Press.
Janvier, P. (1996a). Early vertebrates. Oxford Monographs in Geology and Geophysics, 33, Oxford University Press, Oxford.
Janvier, P. (1996b). The dawn of the vertebrates: characters versus common ascent in current vertebrate phylogenies. Palaeontology, 39, 259-287.
Løvtrup, S. (1977). The Phylogeny of Vertebrata. Wiley, New York.
Stensiö, E. A. (1927). The Devonian and Downtonian vertebrates of Spitsbergen. 1. Family Cephalaspidae. Skrifter om Svalbard og Ishavet, 12, 1-391.
Wang, N. Z. (1991). Two new Silurian galeaspids (Jawless craniates) from Zhejiang province, China, with a discussion of galeaspid-gnathostome relationships.In Early vertebrates and related problems of evolutionary biology (ed. M. M. Chang, Y. H. Liu, and G. R. Zhang), pp. 41-65. Science Press, Beijing.
Information on the Internet
- DigiMorph. The Digital Morphology library is a dynamic archive of information on digital morphology and high-resolution X-ray computed tomography of biological specimens. A National Science Foundation Digital Library at the University of Texas at Austin.
Title Illustrations

| Scientific Name | Selene vomer |
|---|---|
| Specimen Condition | Dead Specimen |
| Body Part | skeleton |
| Copyright | © 1995 Tierney Thys |
About This Page

Muséum National d'Histoire Naturelle Paris, France
Page copyright © 1997
 Page: Tree of Life
Vertebrata. Animals with backbones.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Vertebrata. Animals with backbones.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Citing this page:
Janvier, Philippe. 1997. Vertebrata. Animals with backbones. Version 01 January 1997 (under construction). http://tolweb.org/Vertebrata/14829/1997.01.01 in The Tree of Life Web Project, http://tolweb.org/

