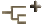Introduction
Gills provide the primary mechanism for respiratory gas exchange in cephalopods as well as a major means of eliminating nitrogenous (ammonia) wastes. Blood is moved through the gills with the aid of the branchial hearts, contractile blood vessels and muscular movements of the gills. Oxygenated blood leaving the gills passes to the muscular ventrical (systemic heart) and is then dispersed via the arterial system to the rest of the animal.
Gill morphology
The gills of Nautilus are distinctive (in addition to having two rather than just one pair of gills) in that the gills lie free in the mantle cavity being attached only at their origin. The gills of coleoids are attached, for varying lengths along one side to the body wall. The general orientation of the gills varies greatly among taxa. For convience we will call the free outer surface of the gill which generally bears the primary efferent vessel, the "top" and the side that is usually attached to the body wall, the "bottom."
The gill has a series of alternating primary lamellae that extend laterally from the gill axis and which form the inner (i. e., medial) and outer (i. e., lateral) demibranchs. The basic structure of the lamellae is similar among the nautiloids and decapodiforms and differs greatly in the octopodiforms.
The branchial gland, where blood hemocyanin is formed, extends along the bottom of the gill as a well-defined structure in coleoids but is scattered throughout the bottom "stem" of the gill in Nautilus in a region (green in the figure below) containing connective and muscle tissue.
Gills of "teuthoids" (myopsids and oegopsids), Vampyroteuthis and incirrate octopods have central holes in the gills that are continuous with the surrounding water between lamellae and, presumably, increase water flow over the gills. These holes align to form a highly perforated "canal" giving the holes the collective name: branchial canal. The branchial canal is absent from nautiloids and sepioids. This feature was one of the primary features used at one time to separately classify sepioids and "teuthoids."

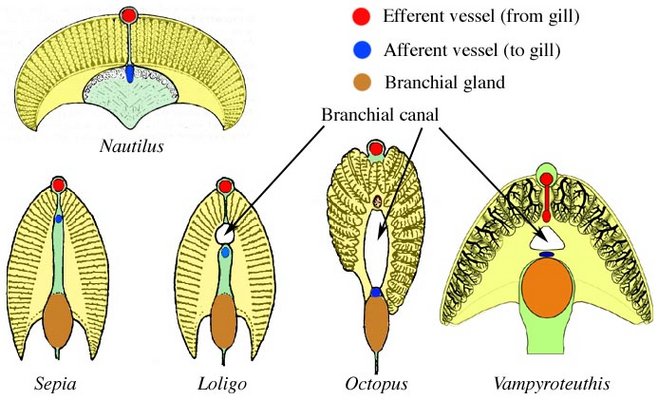
Figure. Cross-sections of gills of various cephalopods to compare their general morphology. Each section shows a primary lamella to either side of the midplane of the gill. incirrate octopods have a marked asymmetry in the size of the two primary lamellae. Asymmetry is also found in oegopsid squids. Drawings, except Vampyroteuthis, modified from Naef (1921-23); Vampyroteuthis modified from Young and Vecchione (2002).
Decapodiform gills
We use Sepia to demonstrate the general form of the gill and the associated blood vessels based on the work of Schipp et al. (1979). Each primary (1°) lamella is folded in a fanlike pattern to form secondary (2°) lamellae that are aligned at right angles to the axis of the 1° lamella. Each 2° lamella, in turn, is folded in a fanlike pattern to from tertiary (3°) lamellae aligned at right angles to the axis of the 2° lamella. Thus the direction of the folding of the 2° and 3° lamellae are at right angles to one another.
The primary (1°) afferent vessel bringing unoxygenated blood to the gill passes near the axis of the gill well above the branchial gland and sends a secondary (2°) branch into each 1° lamellae along the base of the lamella and this 2° afferent vessel in turn sends a branch (3° afferent vessel) over each of the two external edges of the 2° lamellae.
The 1° efferent vessel taking oxygenated blood from the gill passes along the axial plane of the gill but at the top of the gill and receives 2° branches from along the top of each 1° lamella. In turn, a 2° efferent vessel receives 3° vessels that pass directly upward from the 2° lamellae where this lamella passes below it and NOT from the outer edges of the 2° lamella. That is, a 3° efferent vessel runs through the center of a 2° lamella (see drawing at left below and photographs of Histioteuthis below) while a 3° afferent vessels runs along the outer edge of a 2° lamella.
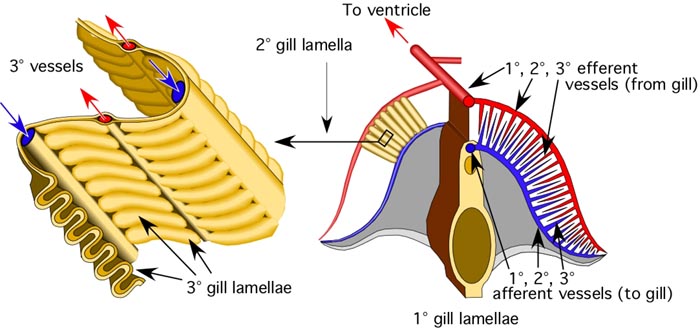
Figure. Diagramatic view of the cross-section of the gill (right) of Sepia officinalis showing the basic circulation of the gill, and an enlargement of a portion of a 2° lamella (left) to show the form of the 2° and 3° lamellae and the locations of their blood vessels. Drawings modified from Schipp, et al. (1979).
Other decapodiforms have similar gills although most oegopsids have thicker 1° lamellae and somewhat asymmetrical gills. In oegopsids the tip of each 1° lamella of the outer demibranch attaches to the base of the gill while the tip of its mate from the inner demibranch extends outward where it is supported by a cartilagenous rod. In Sepia, a cartilagenous rod supports lamellae in both demibranchs.
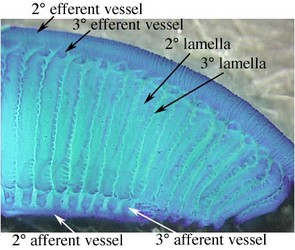
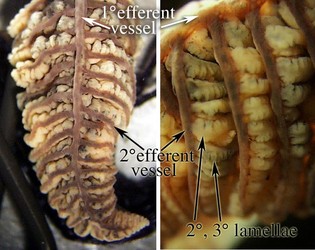
Figure. Left - Side view of a portion of the 1° lamella of Sepia officinalis stained with methylene blue. Right - Top view (left) and side view (right) of portions of the gill of Histioteuthis sp. (Oegopsida). The purple pigmentation of the 1° and 2° efferent vessels is natural. Photographs by R. Young.
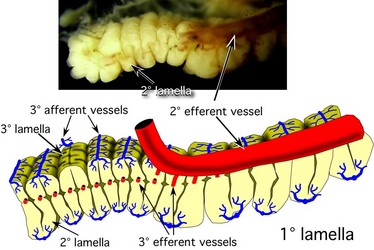
Figure. Top view of a 1° lamella of Histioteuthis sp. with the 1° efferent vessel partially removed to show (1) the fan or Z-fold arrangement of the 2° lamellae and the points where the 3° efferent vessels emerge from the 2° lamellae to join the 2° efferent vessel and (2) the position of the 3° afferent vessels on the external surfaces of the 2° lamellae. Note that the two sides of a 1° lamella are not always equal. Left - Photograph by R. Young. Right - Diagramatic drawing modified from Young and Vecchione (2002).
The photographs below emphasize the fan-fold arrangement of the 2° lamellae which can be unfolded if the constraining ends are cut. A similar effect could be attained with the 3° lamellae.
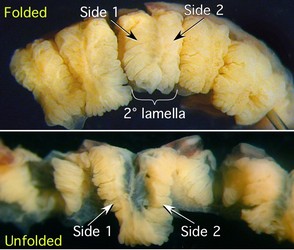
Figure. Side views of a portion of a 1° gill lamella of Histioteuthis. Top - Lamella in its normal folded configuration. Bottom - Membranes at the tip and bottom cut to allow the lamella to be unfolded. The 2° lamella has two (side 1 and side 2) separate members (the respiratory surfaces of the 3° lamellae) on each of its numerous front and back loops. The members of all 2° lamellae are separate from one another except at their extreme ends. The back side of each loop of the 2° lamella has two separated members that are smaller in size than those of the front side. Compare with the diagram of Sepia. Note the subdivisions of the 3° lamellae. Photographs by R. Young.
Octopodiform gills
The octopodiform gill differs dramatically from that of other coleoids. The major difference is in the folding of the gill lamellae. Instead of continuous fan-like folds that form the 2° lamella, the folds have, in effect, become joined in a plane half way between the external edges to form a central "membrane" that separates one side of a 1° lamella from the other side. Similar changes occur in the 3° lamellae where members of an adjacent pair are joined rather than separated by a space.
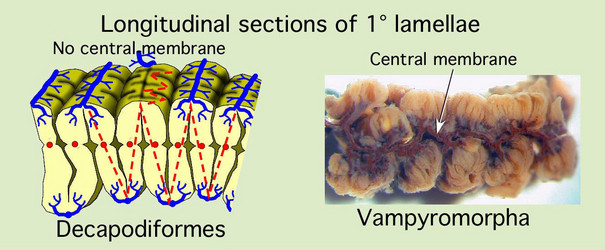
Figure. Left - Longitudinal section through a primary lamella of Histioteuthis sp. (portion of previous drawing) with red-dashed lines that emphasize the two fan-fold patterns. Right - Longitudinal section through a primary lamella of Vampyroteuthis showing the membrane that runs down the midline. This gill had been injected with squid ink several years before the photograph was taken and resulted in the dark appearance of the central membrane. Drawing modified from Young and Vecchione (2002); photograph by R. young
A third major difference between decapodiform gills and octopodiform gills is the form of the 3° lamellae. In decapodiforms, such as Sepia, the 3° lamellae are basically simple folds while in the octopodiformes they are branching folds. Some midwater squids, however, show subdivisions of each 3° lamella and this can become very complex and appear similar to the branching folds of the octopodiforms. However, as far as we are aware, this complex folding never involves branching. That is, if one were to remove an entire 3° lamella from such a squid and unfold it, it would form a flat sheet. A branching lamella cannot be unfolded.
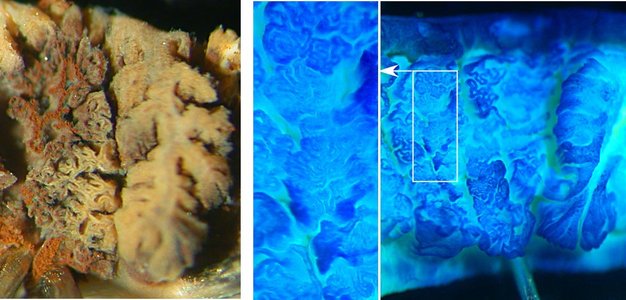
Figure. Left - A top view of two "sets" of 2° lamellae of Vampyroteuthis (octopodiform)with the top of the left set removed with a pair of scissors. The right and left sides of this set are not separate and the folding involves branching. The reddish-brown color is from an old, partial injection of squid ink. Right - A top view of three "sets" of 2° lamellae of Histioteuthis (decapodiform) with the top of the left two sets removed with a pair of scissors. An enlargement of one portion of the photograph is shown in the center. Note the very complex folding but the folds don't branch. The Histioteuthis gill was stained with methylene blue stain. Photographs by R. Young.
Cross-sections of the primary lamellae of the octopodiforms clearly show the distinctive nature of their gills, especially the central membrane and the branching 3° lamellae.
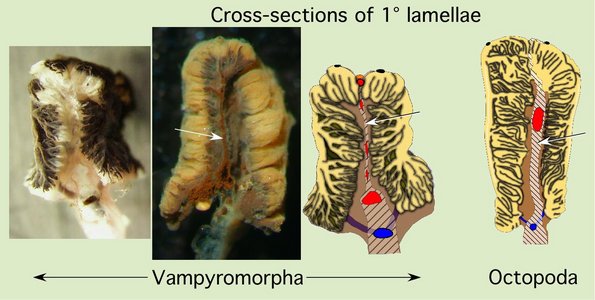
Figure. Left - Two photographic cross-sections of a 1° lamella of Vampyroteuthis (Vampyromorpha) and a diagramatic illustration of the same. The first gill was injected with squid ink through the 1° afferent vessel and shows the ink entering the 2° and 3° lamellae from the outer side. The second gill was injected through the 1° efferent vessel and shows ink (reddish-brown from age) entering more centrally and staining the central membrane. Note the highly branching 3° lamellae. Right - Diagramatic illustation of a cross-section of a 1° lamella of Octopus (Octopoda) with a form very similar to that of Vampyroteuthis. Arrows point to the central membranes in illustrations where it is visible. Photographs by R. Young. Drawings from Young and Vecchione (2002).
Vampyromorpha
The vampire gill is symmetrical with inner and outer demibranchs mirror images of one another. Some aspects of the blood circulation are unique. The afferent circulation is similar to that of Sepia and Octopus with branching vessels passing along the external surfaces of the 2° lamellae. The 1° afferent vessel runs down the axis of the gill just above the branchial gland and just below the branchial canal. Each 2° afferent vessel extends through the tip of the 2° lamella to the outer surface of the lamella where it passes along this outer surface to the top of the gill while sending off branches to the 3° lamellae.
The vampire efferent system, however, is peculiar. The 1° efferent vessel runs along the median line on top of the gill as in most other cephalopod gills. The 2° efferent vessels, however, run along the top of the 1° lamella in only the distal third of the gill. In the proximal two-thirds of the gill, this vessel is absent although the muscular cord that accompanies the vessel distally is still present (see below). For much of the gill, therefore, a secondary efferent drainage is the only system available. This secondary efferent system consists of a large vessel, the "core vessel", that runs down the axis of the gill just above the branchial canal with repeated links to the 1° efferent vessel. The core vessel receives blood from vessels that run along the base of the primary lamellae. The drainage of the 1° lamellae is complex and the septa of these lamellae contain numerous blood spaces that connect with the core vessel. This secondary efferent system is also present in the distal third of the gill along with the secondary efferent vessel which also connects with the blood spaces of the central membrane.
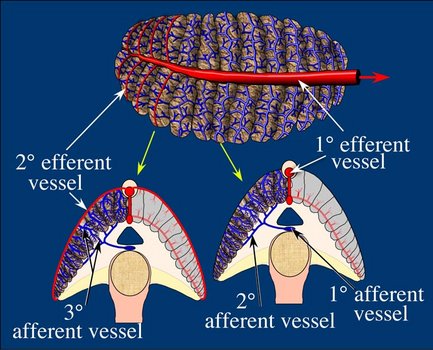
Figure. Blood circulation in the gill of Vampyroteuthis infernalis. Top - A top view of the gill showing efferent vessels in red and afferent vessels in blue. Bottom - Cross-sections of the gill showing circulation to the 1° and 2° lamellae. Yellow arrows show where the cross-sections of the gill were made. Drawing modified from Young and Vecchione (2002).
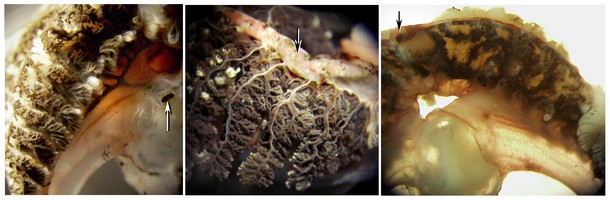
Figure. Various views of ink-injected gills of Vampyroteuthis infernalis. Left - Injection through the 1° afferent vessel (arrow). The 2° afferent vessels can be seen and the ink has entered the outer surfaces of the 2° lamellae. Middle - A more complete injection from the 1° afferent vessel. The 3° lamellae are filled with ink but no ink has reached the 1° efferent vesssel (arrow) which is not directly connected with the ink-filled vessels. Right - The distal region of the gill has been injected through the 1° efferent vessel (arrow) and ink has penetrated through the 2° efferent vessel and the secondary efferent drainage of the central membrane of the 1° lamella. In this picture, the secondary lamellae, which normally obscure the central membrane, have been removed. Photographs by R. Young.
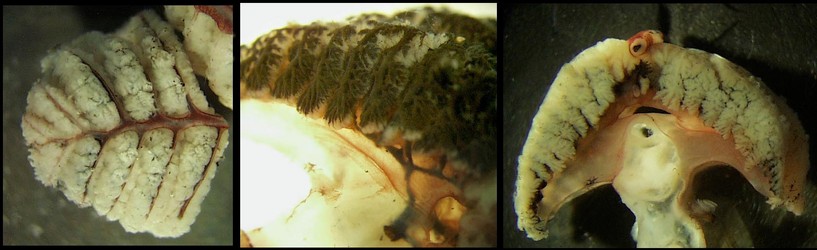
Figure. Left - The distal third of the gill of Vampyroteuthis infernalis showing the 1° and 2° efferent vessels on the top of the gill. Right - Side view of a 1° lamella of a gill injected with ink through the 1° afferent showing the 1°, 2° and 3° vessels and the filling of the 3° lamellae begining at their tops. Photographs by R. Young.
Octopoda: Incirrata
The incirrate gill is asymmetrical in cross-section, somewhat recalling the general morphology of oegopsid squids where the 1° lamellae of the outer demibranch are attached to the gill base but their counterparts of the inner demibranch are free. A branchial canal is present. The afferent circulation is similar to other cephalopods. The 1° efferent vessel runs along the midline of the top of the gill but the 2° efferent vessels run through the center of the 1° lamellae.
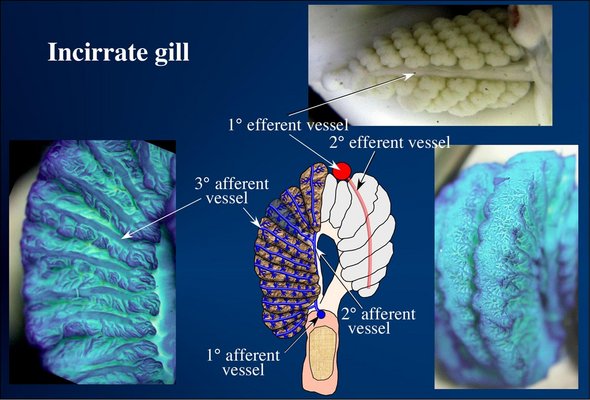
Figure. Blood circulation in the gill of incirrate octopods. Top - A top view of the gill of Octopus cyanea. Bottom left - Portion of a 1° lamella (part of a cross-section through the gill) of Octopus sp. showing the afferent vessel extending along the surface of each 2° lamella and the complex branching of the 3° lamellae. Bottom middle - Drawing of a cross-section through the gill of Octopus. One 2° gill lamella is translucent to show how blood passes from the 2° afferent vessel, through the end of the 2° lamella to emerge on the surface of that lamella. Note the 2° efferent vessel passes through the center of the 1° lamella rather than the outer surface of this lamella. Bottom right - Oblique view of two 1° lamellae of Octopus sp. showing the afferent vessels anastomosing on the outer surface of the lamellae. The blue gills were stained with methylene blue stain. Photographs and drawings modified from Young and Vecchione (2002).
Octopoda: Cirrata
The cirrate gill is quite different from that of incirrates. The cross-section of the gill is symmetrical and neither demibranch is fully attached at its tip to the gill stem. A branchial canal is absent.
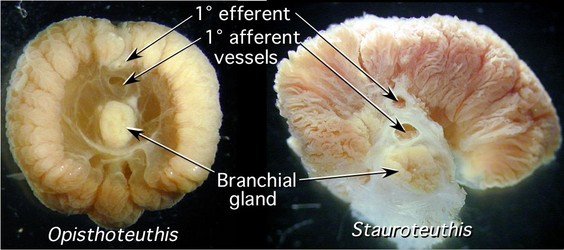
Figure. Cross-sections through the gills of Opisthoteuthis sp and Stauroteuthis syrtensis showing the general structure of the gills. Designation of vessels as afferent or efferent is based on their relative position not on definitive information. Photographs by R. Young.
The general external form of the cirrate gill varies greatly among taxa and in some the primary lamellae are difficult to recognize (Stauroteuthis).
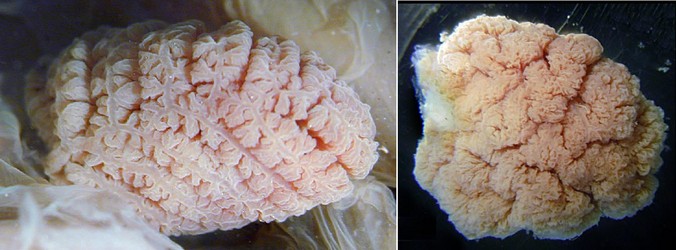
Figure. Top views of the gills of Cirroteuthis muelleri (left) and Stauroteuthis syrtensis (right) showing the very different appearance. Photographs by R. Young.
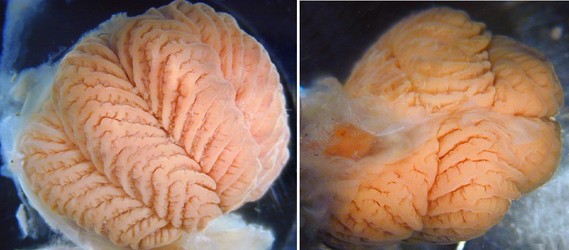
Figure. Anterior (left) and posterior (right) views of the gill of Cirroctopus glacialis. From the anterior view, the gills has the “half-orange” appearance while from the posterior view, the gill has a more normal appearance with 1° lamellae of either side of the gill. Note the characteristic (for Cirroctopus) “Z” formation in the midline of the 1° lamellae where the 2° lamellae merge. Photographs by R. Young.
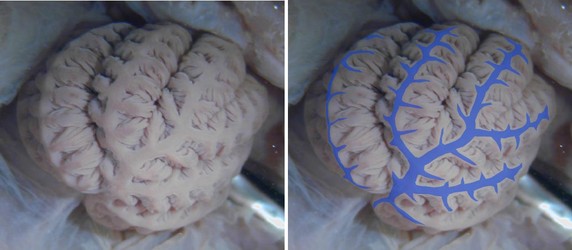
Figure. Left - Top view of the gill of Grimpoteuthis tuftsi. Right - Same photograph with the afferent vessels indicated by blue coloration. Superficially the gill resembles that of a decapodiform with a large central vessel and branching vessels down the top of each 1° lamella; however, these vessels in decapodiforms are efferent not afferent. The anastomosing of the outer afferent blood vessels seen in Octopus, has apparently been accentuated in cirrates. This arrangement is also easily seen in the photograph of Cirroteuthis muelleri found higher on this page. Photograph by R. Young.