Cephalopod beaks have been studied for many years primarily for their value in cephalopod taxonomy and, especially, in identifying cephalopods from stomach contents (e.g. Clarke, 1986). Some gaps, however, remain in our understanding of beak stucture, morphology and evolution; some of these are addressed here in this and planned reviews.
Basic structure of the cephalopod beaks
Although in many cephalopods the upper (dorsal) and lower (ventral) beaks look very different, their basic structure is virtually the same:
1. Each beak consists of two primary components, an outer one (the outer beak or "hood") and an inner one (the inner beak or "wall"). Each of these components are roughly shaped like a half-cone.
2. Each beak also consists of two secondary components, the Bridge and the Shoulder. The Bridge is a long, slender strap that joins together the two primary components (i.e. hood and wall). The Bridge extends from the beak tip posteriorly and fuses to the hood along its outer edge and to the wall along its inner edge.

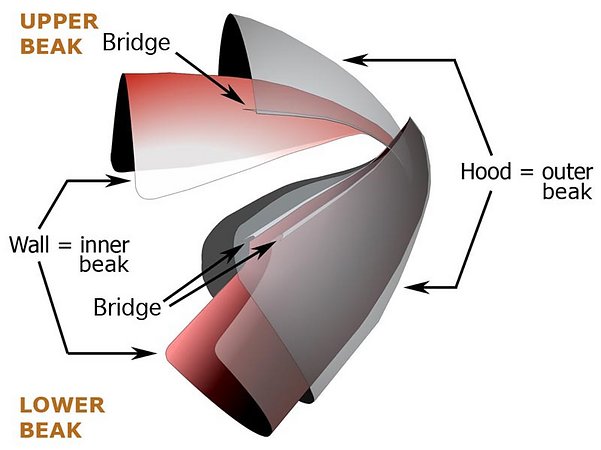
The Shoulder plays a more complex, and not well understood, role but also participates in joining the "hood", "wall" and "bridge". The drawing below shows the position of the shoulder but oversimplifies its shape.

While the basic structure of the upper and lower beaks are the same, there are some distinct differences between them that result from their different roles in the biting process with the upper beak being the inside component of the bite and the lower beak the outside component; that is, the upper beak must bite inside the lower beak.
As a result of the way the beaks bite the upper beak has:
- the jaw edge formed at the fusion of the Hood and Bridge (see above drawings);
- the shoulder fused to the Wall and Bridge (see above drawings).
In contrast the lower beak has:
- the jaw edge formed at the fusion of the Wall and Bridge (see above drawings);
- the shoulder fused to the Hood and Bridge (see above drawings).
The beaks have no rigid hinge which, at first look seems peculiar and, perhaps, an evolutionary failure. However the way the beaks operate preclude the use of a fixed hinge. The beaks cut prey somewhat like a pair of scissors working in reverse. That is, the cut begins at the tip of the scissors and progresses toward the base. To accomplish this, the beaks have a muscular articulation (i.e., a flexible joint) that allows considerable movement of the pivot area (Uyeno and Kier, 2005) and this allows the two cutting edges of the upper beak to move along the two cutting edges of the lower beak as the upper beak descends into the confines of the lower beak.
Comparison of Vampyroteuthis infernalis beaks and decapodiform beaks


Figure. Top - Vampyroteuthis infernalis. Bottom - Asperoteuthis acanthoderma, our representative of the more primitive decapodiform beaks. Left - Upper beaks. Right - Lower beaks. Photographs by R. Young. (3D)
In describing beaks of V. infernalis and A. acanthoderma we will use a more detailed terminology. However, we will retain the use of Hood to refer to the outer beak and Wall to refer to the inner beak. An overview of beak terminology can be found here. Briefly (see photographs below), the Wall consists two regions: a lateral region ob either side called the Lateral Wall and a top (= dorsal) region called the Crest. (In life, the lower mandible is situated upside down relative to the upper mandible which means that the Crest is dorsal on one and ventral on the other. This can get very confusing. As a result we apply dorsal, ventral, anterior, posterior terms the same on both mandibles based on their standard orientation in the figures above) The Hood differs between upper and lower beaks. Both beaks have an anterior region called the Rostrum and a Posterior region called the Posterior Hood. In addition the Hood of the lower beak has an extension called the Wing (see Lower Beak below). The Shoulder consists of an Outer Part with a modified, pigmented region (Shoulder Blade) adjacent to the embedded continuation of the jaw edge (= Jaw Edge Extension), and a cartilagenous-like, unpigmented region that forms much of the bulk of the shoulder (Hyaline Matriix, a term we borrow from "hyaline cartilage" of vertebrates). The mostly unpigmented material of the young beak (ie, most of the Wall and Hood) has the same appearance as Hyaline Matrix but is termed hyaline material and the two differ only, as far as we know, in thickness and location. The Hyaline Matrix is found only in the shoulder and onto the lateral areas of the Palate (the Bridge). Hyaline material can be found virtually everywhere else.
Upper Beak
The following pictures shows the more detailed terminology and the very similar appearance of the V. infernalis and the A. acanthoderma (Chiroteuthidae) beaks.
The Hood and Lateral Wall are easily recognized (photographs below). The Shoulder Blade is a distinct, pigmented structure in the upper beaks of both species. It is the uppermost (dorsal) part of the Outer Part of the Shoulder and, often, extends anteriorly slightly beyond the rest of the Outer Part. It abuts the Jaw-edge Extension. Much of the Inner Part of the shoulder, in both species, is the anterior region of the Lateral Wall which extends well anterior to the Shoulder Blade in these photographs.
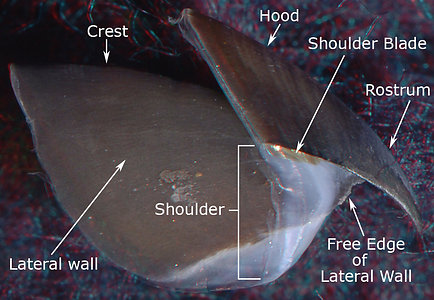
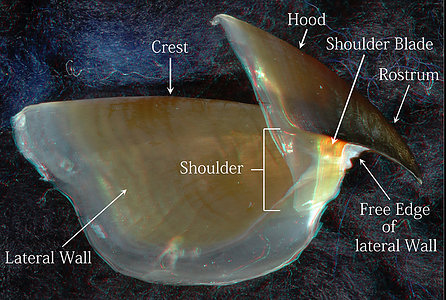
Figure. Side views of the upper beaks showing the basic terminology. Left - V. infernalis. Right - A. acanthoderma. (3D)
The anterior Free Edge of the Lateral Wall, extends onto the Pallet as a low, sharp ridge that defines the inner edge of the Bridge (photographs below). The Bridge anteriorly is covered more completely by the Hyaline Matrix in A. acanthoderma than in V. infernalis but can still be seen, in the former, through the translucent Hyaline Matrix. In the photograph of V. infernalis below, the Bridge can be seen throughout virtually its entire length, even where it is covered by the translucent Hyaline Matrix that fills most of the Middle Part of the Shoulder of both species. The Hyaline Matrix looks much like vertebrate cartilage but is very different. The entire beak is a secreted structure and contains no cells. The Hyaline Matrix is well-hydrated, softer and a more flexible material that the stronger, tougher pigmented (i.e., sclerotized) portions of the beak. In the sclerotized portions, much of the water has been extruded and the high protein content has formed numerous cross-linkages within a chitinous scaffold (Miserez et al., 2010).


Figure. Left - Oral view of the upper beak of V. infernalis. Right - Oblique view of the anterior half of the upper beak of A. acanthoderma. (3D)


Figure. Posterior views of the upper beaks. Left - V. infernalis. Right - A. acanthoderma. (3D)
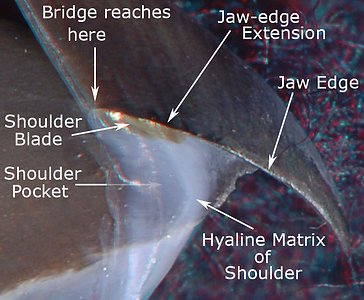
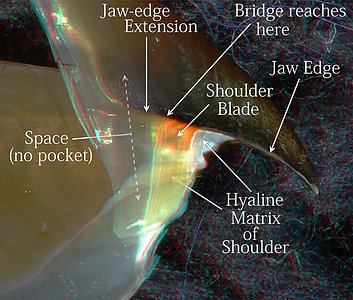
Figure. Side view of the Shoulder regions of the upper beaks. Left - V. infernalis. Note that the arrow marked "Shoulder Pocket" is dimmed toward the arrow point to indicate a transparent Outer Part of the pocket that isn't obvious without 3D glasses. Right - A. acanthoderma. Note the Outer Part of the Shoulder below the Shoulder Blade has developed a slight pigmentation at this size (300 mm ML). Photograph by R. Young. (3D)
Lower beak
The lower beak of V. infernalis has an extremely large Hood which makes the beak easy to recognize but also makes it look very different from A. acanthoderma. The small Hood in the latter provides clear differentiation of the Wing. The Jaw Edge is relatively long in V. infernalis and its Shoulder Blade, while in the same relative position as that of A. acanthoderma (i.e., it extends posteriorly from the Jaw Angle along the Jaw-edge Extension), is relatively large and partially obscured by a white covering of hyaline material. As in the upper mandibles, both the Shoulder Blade and the Jaw-edge Extension in V. infernalis reach virtually to the anteroventral end of the Lateral Wall (see below) but fall well short of this in A. acanthoderma (see well below). In contrast to the upper mandibles, the Hyaline Matrix of both taxa lies "above" (anterodorsal to) the Bridge rather than "beneath" (posteroventral to) the Bridge as in the upper beak (of course, if you turn the lower beak upside-down then the Hyaline Matrix would be in the same relative position as in the upper beak). The Free Edge of the Hood is extensive in the shoulder region and extends to the rostral tip in V. infernalis. In A. acanthoderma, the free edge of the Hood, while protruding less, occupies virtually the same position as in V. infernalis.
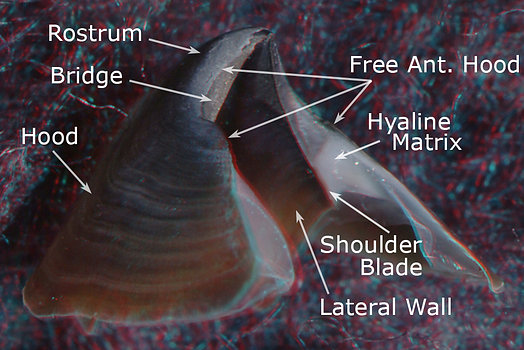
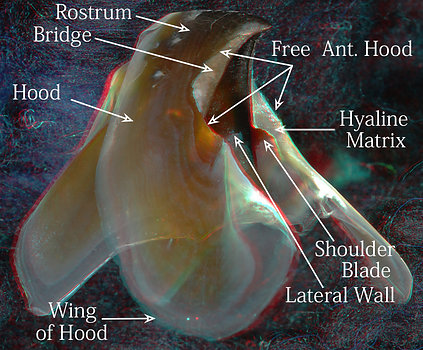
Figure. Left - Side-oblique view of the lower beak of V. infernalis, showing the basic features (3D). Right - Side-oblique view of the lower beak of Asperoteuthis acanthoderma.
The posterior view of V. infernalis (photographs below) allows one to nearly see the full extension of the Bridge. To show that the Bridge of V. infernalis is virtually a flat strap that joins the lateral wall and the Hood, we have made cuts through two beaks at different places in the Shoulder. One (middle photograph below) was cut just anterior to the Shoulder where the Free Edge of the Hood rises only slightly above the Bridge and shows the Bridge connecting the Jaw Edge of the Lateral Wall with the Hood. The other (right photograph) was cut through the middle of the Shoulder containing much Hyaline Matrix where the Hood rises far above the Bridge, lateral to the Hyaline Matrix, and shows the Bridge connecting the Jaw-edge Extension with the Hood. The Jaw-edge Extension and the Bridge in the lower beak of V. infernalis extends virtually to the back end of the Lateral Wall (as mentioned earlier). The clear separation of the Lateral Wall and Shoulder by the Jaw-edge Extension in V. infernalis, suggests that the inner wall of the Shoulder is not a part of the Lateral Wall. As in the upper beak a Shoulder Pocket (see middle image, not labeled) is present in V. infernalis.
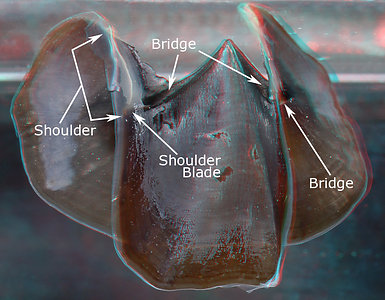
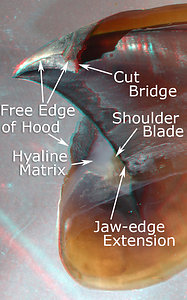

Figure. Various views of the lower beak of V. infernalis. Left -Posterior view, intact beak. Middle - Oral-lateral view with most of the right side removed. Right - Posterolateral view with much of the right side removed. (3D)

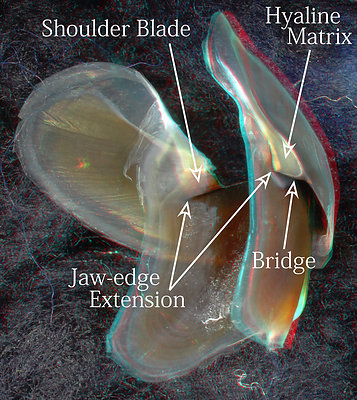
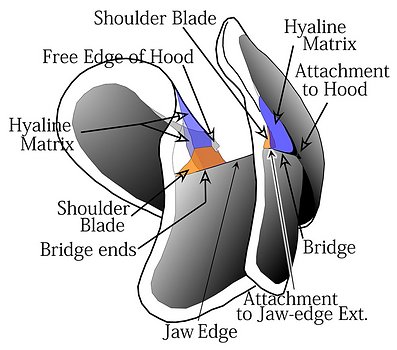
Figure. Various views of the lower beak of A. acanthoderma. Top left - Oral view, photograph (3D). Top right - Posterolateral view, photograph (3D). Bottom - Posterolateral view, drawing (2D).
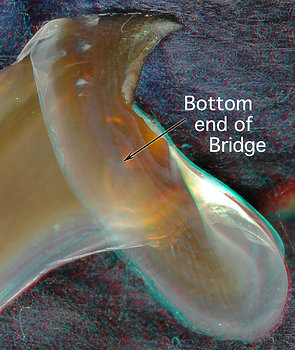

Figure. Two views of the lower beak of A. acanthoderma. Left - Side view of front region of beak. Right - Back-oblique view of central region of beak. (3D)
Comments
The upper beak of A. acanthoderma is very similar to that of V. infernalis, except for its lesser posterior reach of the Bridge, and to a lesser extend, the Jaw-edge Extension.
The lower beak of A. acanthoderma differs greatly in appearance from that of V. infernalis due its much smaller hood and wings. In addition, the lower beak of A. acanthoderma, as in the upper beak, has a lesser extent of the Bridge and Jaw-edge Extension than does V. infernalis.
The Free Edge of the Hood at the front of the lower beak in A. acanthoderma is strikingly similar to that of V. infernalis. In A. acanthoderma, this feature appears, in the region of the Shoulder, rather late in ontogeny, as a structure that extends over and slightly beyond the Wing Fold. A Wing Fold is not recognizable in V. infernalis. The Wing Fold in decapodiforms, however, is formed first (during development) by a thick layer of Hyaline Matrix that lies over (i.e., anterodorsal to) the Bridge, suggesting that the origin of the Wing Fold is from the latter (i.e., the Middle Part of the Shoulder) and not from the Wing itself.
A major difference in both mandibles between V. infernalis and A. acanthoderma is the fate, during growth, of the posterior end of the Bridge and its associated Hyaline Matrix.
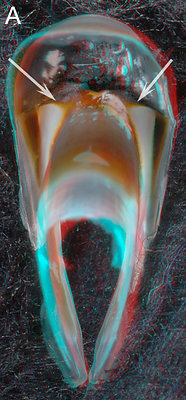

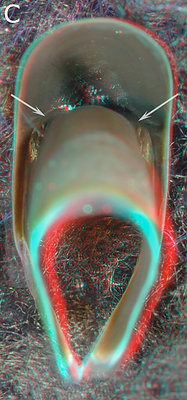

Figure. A - Aperoteuthis acanthoderma, female, 300 mm ML, 5.1 URL. B - A. acanthoderma, presumably male, 250 ? mm ML, 7.0 mm URL. C - Galiteuthis pacifica, Male, 333 mm ML, 4.6 mm URL. D - Vampyroteuthis infernalis - ? mm ML, 6.2 mm URL.
Figs. A - C show the changes in the pigmentation of the Shoulder with age. Unfortunately we didn't have a series from one species so we have presented a composite series of two closely related species. The series represents a change in extent of beak pigmentation rather than in size (size is affected by species and sex differences). Fig. A is a relative young beak (compare to posterior upper beak views earlier on this page between young Asperoteuthis acanthoderma and V. infernalis). The arrows point to the Bridge; the light-colored area below it is the Hyaline Matrix. In Fig. B the Hyaline Matrix of the posterior Shoulder is more heavily pigmented. Fig. C shows the posterior Shoulder fully pigmented. Fig. D shows the right side of the largest beak of a V. infernalis that we have seen, which is nearly three times the length of the posterior view shown earlier on this page, yet the Bridge looks no different. The anterior end of the beak and Hood have been removed which provides a clear view of the Bridge. Arrows - All point to the Bridge.
We suspect that the Hyaline Matrix never becomes pigmented in V. infernalis. Even if it did, with little Hyaline Matrix directly in content with the posteroventral end of the Bridge, the posterior Shoulder would probably not look much different than seen in Fig. D above. This is clearly not the case in the three cranchiids seen above. The change from a bridge with a free-standing end (attached on all sides except the posterior end) to a fully pigmented, solid posterior Shoulder in decapodiforms involves changes in the structure of the Bridge and the Lateral Wall. It often difficult to sort out which, or both, of the two are mostly responsible. As a result we call the new structure the Lateral-wall Bridge Continuum (= LW-B Continuum). The means by which this occurs helps define the three major types of Upper Beaks found in the Decapodiformes and is critical to understanding their evolution. The example above emphasizes the upper beak but the lower beak has the same problem but the decapodiform solution has yet to be investigated.
In evolutionary terms, V. infernalis represents the outgroup to the Decapodiformes so that the characters shared in common with A. acanthoderms (and those species it represents) can be considered primitive (plesiomorphic) characters in the Decapodiformes. The establishment of such characters is essential if we are to understand how the beaks evolved among members of the Decapodiformes. That is we must be able to recognize new (apomorphic) features that have risen among decapodiforms. Characters that these two taxa don't share could represent apmorphic features of the Vampyromorpha or features that are plesiomorphic in the Coleoidea. The latter possibility would also provide insight into the evolution of decapodiform beaks. This is the primary objective of this page: To point out the features shared and not shared among these two taxa.


