




Aspredinidae
Banjo Catfishes
John P. Friel

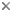
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxmodified from Friel (1994).
Notes about Terminal Taxa: Taxa whose names appear in quotes on this Tree of Life page are undescribed. In accord with articles 8 and 9 of the third edition of the International Code of Zoological Nomenclature (ICZN, 1985), I do not issue this page for permanent scientific record. This page is designated an unavailable work for nomenclatural purposes. All names for new genera and species are provisional and will become available when my manuscript describing the type specimens is published in an official work such as a peer reviewed journal.
Introduction
The Aspredinidae are known as banjo catfishes due to their overall body shape, a depressed head and slender caudal peduncle which somewhat resembles a banjo. They occur throughout the tropical rivers of South America. Local names for aspredinids include "banjaman" or "banjo-man" (Guyana), "croncron" (French Guiana), "rabeca" (Brazil), and "guitarillo" (Venezuela). Banjo catfishes maybe found in habitats ranging from shallow backwaters to deep river channels to tidal estuaries. In general, most species are cryptically pigmented, benthic and sluggish unless disturbed. Many are semi-fossorial, during the day often resting just beneath the substrate surface.
Approximately 6O extant species of banjo catfishes have been described. A large proportion of these species are now considered subjective junior synonyms of earlier described species. As currently recognized the family contains approximately 35 nominal species placed in 13 genera. In addition there are several undescribed species mostly in the genus Bunocephalus. Despite the relatively small number of species in this family as compared to other catfish families, aspredinids are quite diverse in their morphology. They range from miniature armored species such as Hoplomyzon papillatus, less than 20 mm in length, to large elongate species such as Aspredo aspredo, reaching up to 380 mm in length.
Aspredinids are a highly derived group of catfishes and display some very unusual features. Their skin is completely keratinized and covered with tubercles. Periodically the entire outer layer of skin is shed just like that of amphibians and reptiles (Friel, 1989).
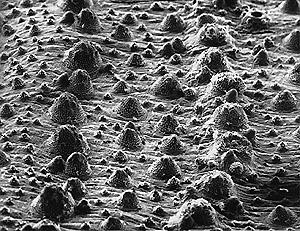 image info
image info
This SEM micrograph shows rows of enlarged tubercles which run longitudinally along the bodies of aspredinids. The light spots covering the tubercles are unicellular keratinized processes called unculi (Roberts, 1982). The horizontal field of view of this image is 1.68 mm.
While aspredinids can swim by typical undulatory movements, they can also use jets of water thrust from their opercular openings to skip along the substrate. When agitated, some species produce audible stridulatory sounds by repeatedly abducting and adducting their pectoral spines.
Very little is known about the general ecology of aspredinids. Based on little published work and personal observation, most aspredinids appear to be generalized omnivores and their stomachs often contain aquatic invertebrates, terrestrial insects and organic debris. One notable exception are members of the genus Amaralia. Based on stomach contents they appear to feed on the eggs of other catfishes (Friel, 1992).
Few specifics are known about reproduction of aspredinids. Parental care is known with certainty in one clade which contains Pterobunocephalus, Platystacus, Aspredo, and Aspredinichthys. Females of this clade carry their developing embryos attached to the ventral surface of their bodies. In Pterobunocephalus, the eggs are directly attached to the body whereas in Platystacus, Aspredo, and Aspredinichthys they are attached to fleshy stalks, called cotylephores, which grow out from the female (Friel, 1994). These develop seasonally and may function in the exchange of materials between the mother and her developing embryos (Wetzel, Wourms, & Friel, 1997).
Discussion of Phylogenetic Relationships
As already mentioned, aspredinids are highly derived catfishes and their monophyly is well supported by many apomorphies. Some of these including: laminar processes of the pterotics directed laterally; vomer absent; mandibular lateralis canal does not enter lower jaw; opercle "L" shaped, resembles a branchiostegal ray; opercular apertures reduced to ventral slits; 5 or fewer branchiostegal rays; dorsal lamina of Weberian complex contacts dorsal surface of body; hemal canal forms de novo by vertebra 7 without a transformation series of rib parapophyses; abdominal vertebral with peg and socket articulations; parapophyses for ribs reduced or absent; 10 or fewer principal caudal-fin rays; expanded bases on outermost caudal-fin rays; muscles on the ventral surface of the pectoral girdle highly reduced or absent; basipterygia without anterior arms; posterior cartilage of basipterygia reduced; mental barbel bases reduced; rows of unculiferous tubercle present on body; and loss of alarm cells & fright reaction. For a complete list of synapomorphies see Friel (1994).
Friel's (1994) phylogenetic revision of the aspredinids revealed that the traditionally recognized subfamily Aspredininae is not the sister group to all other Aspredinidae but is nested higher up in the phylogeny of aspredinids. Furthermore, the subfamily Bunocephalinae sensu Myers (1960), tribe Bunocephalini sensu Myers (1960), Dysichthys sensu Mees (1988, 1989) and Aspredo sensu Mees (1987) are paraphyletic taxa. Major taxonomic and nomenclatural changes are necessitated as a result of this new phylogeny. Species originally placed in Bunocephalus Kner, 1855 and recently transferred to Dysichthys Cope, 1874 by Mees (1988, 1989) are reassigned to "Pseudobunocephalus" new genus, Pterobunocephalus Fowler, 1943, and Bunocephalus Kner, 1855. In addition, several genera are synonymized. Petacara B?hlke, 1959 is a junior subjective synonym of Pterobunocephalus Fowler, 1943; Dysichthys Cope, 1874 is a junior subjective synonym of Bunocephalus Kner, 1855; and Bunocephalichthys Bleeker 1858 and Agmus Eigenmann, 1910 are junior objective synonyms of Bunocephalus Kner, 1855.
The phylogenetic relationship of the Aspredinidae to other catfishes remains controversial. Prior ideas on relationships are briefly summarized. G?nther (1864) first suggested a relationship between the Aspredinidae and a clade containing the Neotropical loricarioids and the Asian Sisoridae. A relationship with loricarioid catfishes was also suggested by Chardon (1968). However Baskin (1972) and Howes (1983) both reviewed Chardon's evidence and concluded that the Aspredinidae are not closely related to loricarioids. Ferraris (1989) suggested that the Asian Akysidae are the sister group to the Aspredinidae. Mo (1991) placed the Aspredinidae either basal to or in a polytomy with a clade containing the Afro-Asian Clariidae, the African Amphiliidae, Neotropical loricarioids, and Asian sisoroids (Amblycipitidae, Akysidae and Sisoridae). Pinna (1993) placed the Aspredinidae in a polytomy with Amblycipitidae, Akysidae, Sisoridae, Amphiliidae and loricarioids. Chen (1994) placed the Aspredinidae as the sister group to Asian sisoroids. More recently, Pinna (1996) now places the Aspredinidae within the currently recognized Asian Sisoridae. Friel (unpublished) reanalyzes all prior evidence presented by others along with new character information and finds two equally parsimonious placements for the Aspredinidae. The sister group to the Aspredinidae are either the Asian sisoroid catfishes as has been suggested by others or the doradoid catfishes (the African Mochokidae, and the Neotropical Doradidae, Centromochlidae and Auchenipteridae).
Geographic Distribution
This Neotropical family of catfishes is found throughout the tropical rivers of South America (Magdalena, Orinoco, Amazon, S?o Francisco and Paraguay-Paran?), a few rivers west of the Andes (Atrato, San Juan, and Patia) and in brackish and marine waters between the Orinoco and Amazon River deltas.
 image info
image info
Key to Genera
- 1a. > 50 anal-fin rays; caudal peduncle with a continuous bony dorsal ridge...2
- 1b. < 20 anal-fin rays; caudal peduncle without a continuous bony dorsal ridge...4
- 2a. 9 principal caudal-fin rays; tubercle rows well developed...Platystacus
- 2b. 10 principal caudal-fin rays; small barbel at base of maxillary barbel present...3
- 3a. Maxillary barbel completely adnate, enlarged humeral process, tubercle rows on body highly reduced; 7 pectoral-fin rays...Aspredo
- 3b. Maxillary barbel not adnate; additional pairs of barbels on ventral surface of body; 8 pectoral-fin rays; antorbitals and mesethmoid with hook-like processes...Aspredinichthys
- 4a. Lower lip equal to upper lip; outermost caudal fin rays shortened; anal and dorsal fins not adnate; lateral line truncated at level of dorsal fin...5
- 4b. Lower lip subequal to upper lip; anal and dorsal fins partially or totally adnate; lateral line complete...6
- 5a. Rigid, locking dorsal spine and spinelet present...Acanthobunocephalus
- 5b. Dorsal spine flexible; no dorsal spinelet..."Pseudobunocephalus" new genus
- 6a. No obvious bony plates on body...7
- 6b. Dorsal, ventral and lateral series of bony plates present on body...10
- 7a. Lower lip with numerous papillae; no premaxillary teeth...Xyliphius
- 7b. Lower lip without papillae; premaxillary teeth present...8
- 8a. 2-3 dorsal-fin rays; caudal peduncle deep, laterally compressed...Amaralia
- 8b. >4 dorsal-fin rays; caudal peduncle slender, not laterally compressed...9
- 9a. Head and body extremely depressed; skull ornamentation highly reduced or absent, anal-fin with 10 - 20 rays...Pterobunocephalus
- 9b. Head and body deep or moderately depressed; skull ornamentation typically developed; anal fin with 5 - 10 rays...Bunocephalus
- 10a. Papillae present on upper lip; maxillary barbels completely adnate; dentary teeth present; 3 - 4 sets of paired pre-anal-fin plates...Hoplomyzon
- 10b. No papillae present on upper lip; maxillary barbels on slightly adnate; dentary teeth absent; 1 - 2 sets of paired pre-anal-fin plates...11
- 11a. Eyes present; heavily pigmented...12
- 11b. Eyes absent; pigmentation reduced...Micromyzon
- 12a. One set of paired pre-anal-fin plates; pectoral spines only slightly longer than first branched ray; skull ornamentation well developed...Dupouyichthys
- 12b. Two sets of paired pre-anal-fin plates, pectoral spines much longer than first branched ray; skull ornamentation reduced...Ernstichthys
List of Synonymies
| Original Description Name | Current Placement |
| Acanthobunocephalus nicoi Friel, 1995 | Acanthobunocephalus nicoi |
| Agmus lyriformis Eigenmann, 1912b | Bunocephalus verrucosus |
| Aspredinichthys filamentosus Valenciennes, 1840 | Aspredinichthys filamentosus |
| Aspredinichthys tibicen Valenciennes, 1840 | Aspredinichthys tibicen |
| Aspredo aspredo Linnaeus, 1758 | Aspredo aspredo |
| Aspredo batrachus Gronovius, 1854 | Aspredo aspredo |
| Aspredo sexcirrhis Valenciennes, 1840 | Platystacus cotylephorus |
| Aspredo sicuephorus Bleeker, 1858 | Aspredo aspredo |
| Aspredo spectrum Gronovius, 1854 | Platystacus cotylephorus |
| Bunocephalus albifasciatus Fowler, 1943 | Pterobunocephalus depressus |
| Bunocephalus aleuropsis Cope, 1870 | Bunocephalus aleuropsis |
| Bunocephalus amaurus Eigenmann, 1912b | Bunocephalus amaurus |
| Bunocephalus amaurus aloike Hoedeman, 1961 | Bunocephalus amaurus |
| Bunocephalus amaurus sipaliwini Hoedeman, 1961 | Bunocephalus amaurus |
| Bunocephalus bicolor Steindachner, 1882 | Bunocephalus coracoideus |
| Bunocephalus bifidus Eigenmann, 1942 | "Pseudobunocephalus" bifidus |
| Bunocephalus "boliviensis" Ma, 1977 | "Pseudobunocephalus" amazonicus |
| Bunocephalus carvalhoi Miranda Ribeiro, 1944 | "Pseudobunocephalus" iheringii |
| Bunocephalus chamaizelus Eigenmann, 1912b | Bunocephalus chamaizelus |
| Bunocephalus colombianus Eigenmann, 1912a | Bunocephalus colombianus |
| Bunocephalus coracoideus Cope, 1874 | Bunocephalus coracoideus |
| Bunocephalus depressus Haseman, 1911 | Pterobunocephalus depressus |
| Bunocephalus dolichurus Delsman, 1941 | Pterobunocephalus dolichurus |
| Bunocephalus doriae Boulenger, 1902 | Bunocephalus doriae |
| Bunocephalus "dorsolineatus" Ma, 1977 | "Pseudobunocephalus" sp.? |
| Bunocephalus gronovii Bleeker, 1858 | Bunocephalus verrucosus |
| Bunocephalus haggini Eigenmann & Allen, 1942 | Bunocephalus coracoideus |
| Bunocephalus hypsiurus Kner, 1855 | Amaralia hypsiura |
| Bunocephalus iheringii Boulenger, 1891 | "Pseudobunocephalus" iheringii |
| Bunocephalus knerii Steindachner, 1882 | Bunocephalus knerii |
| Bunocephalus larai von Ihering, 1930 | Bunocephalus larai |
| Bunocephalus melas Cope, 1874 | Bunocephalus aleuropsis |
| Bunocephalus minutus G?ntert, 1942 | "Pseudobunocephalus" iheringii |
| Bunocephalus retropinnis Eigenmann, 1942 | Bunocephalus doriae |
| Bunocephalus rugosus Eigenmann & Kennedy, 1903 | "Pseudobunocephalus" rugosus |
| Bunocephalus salathei Myers, 1927 | "Pseudobunocephlaus" iheringii |
| Bunocephalus scabriceps Eigenmann & Eigenmann, 1889 | Bunocephalus verrucosus |
| Bunocephalus "spieleri" Ma, 1977 | "Pseudobunocephalus" sp.? |
| Cotylephorus blochii Swainson, 1838 | Platystacus cotylephorus |
| Dupouyichthys sapito Schultz, 1944 | Dupouyichthys sapito |
| Dysichthys amazonicus Mees, 1989 | "Pseudobunocephalus" amazonicus |
| Dysichthys australe Eigenmann & Ward, 1907 | "Pseudobunocephalus" rugosus |
| Dysichthys quadriradiatus Mees, 1989 | "Pseudobunocephalus" quadriradiatus |
| Ernstichthys anduzei Fern?ndez-Y?pez, 1953 | Ernstichthys anduzei |
| Ernstichthys intonsus Stewart, 1985 | Ernstichthys intonsus |
| Hoplomyzon megistus Orc?s, 1961 | Ernstichthys megistus |
| Hoplomyzon atrizona Myers, 1942 | Hoplomyzon atrizona |
| Hoplomyzon atrizona petroleus Schultz, 1944 | Hoplomyzon atrizona |
| Hoplomyzon papillatus Stewart, 1985 | Hoplomyzon papillatus |
| Hoplomyzon sexpapillostoma Taphorn & Marrero, 1990 | Hoplomyzon sexpapilostoma |
| Micromyzon akamai Friel & Lundberg, 1996 | Micromyzon akamai |
| Platystacus cotylephorus Bloch, 1794 | Platystacus cotylephorus |
| Platystacus laevis Bloch, 1794 | Aspredo aspredo |
| Platystacus nematophorus Bleeker, 1862 | Platystacus cotylephorus |
| Platystacus verrucosus Bloch, 1794 | Bunocephalus verrucosus |
| "Pseudobuncephalus lundbergi" n. sp | "Pseudobuncephalus lundbergi" n. sp. |
| Siluris hexdactylus La Cepede, 1803 | Platystacus cotylephorus |
| Siluris verrucosus Walbaum, 1792 | Bunocephalus verrucosus |
| Xyliphius barbatus de Ar?mburu & Ar?mburu, 1962 | Xyliphius barbatus |
| Xyliphius kryptos Taphorn & Lilyestrom, 1983 | Xyliphius kryptos |
| Xyliphius lepturus Orc?s, 1962 | Xyliphius lepturus |
| Xyliphius lombarderoi Risso & Risso, 1964 | Xyliphius lombarderoi |
| Xyliphius magdalenae Eigenmann, 1912a | Xyliphius magdalenae |
| Xyliphius melanopterus Orc?s, 1962 | Xyliphius melanopterus |
References
Baskin, J. N. 1972. Structure and relationships of the Trichomycteridae. Ph.D. thesis, City University of New York, XXI + 389 pp.
Böhlke, J. E. 1959. Results of the Catherwood Foundation Peruvian Amazon expedition. Petacara, a new genus for the bunocephalid catfish, Bunocephalus dolichurus Delsman. Notulae Naturae, 318: 1-6.
Chardon, M. 1968. Anatomie comparée de l'appareil de Weber et des structures connexes chez les Siluriformes. Annales de Muse Royale de l'Afrique Centrale, Ser. 8, Sciences Zoologiques, 169: 1-227.
Chen, X. 1994. Phylogenetic studies of the amblycipitid catfishes (Teleostei, Siluriformes) with species accounts. Ph.D. thesis, Duke University, XX + 471 pp.
Cope, E. D. 1874. On some Batrachia and Nematognathi brought back from the upper Amazon by Prof. Orton. Proceedings of the Academy of Natural Sciences of Philadelphia, 120-137.
Eigenmann, C. H. 1910. Catalogue of the fresh-water fishes of tropical and South America. Reports of the Princeton University Expeditions to Patagonia, 1896-1899, 3(2):375-484.
Ferraris, C. 1989. On the interrelationships between the Aspredinidae and the Akysidae (Ostariophysi, Siluriformes). Abstracts of the 1989 Annual Meeting of the American Society of Ichthyologists and Herpetologists at San Francisco StateUniversity, p. 86.
Fowler, H. W. 1943. Zoological results of the second Bolivian expedition for the Academy of Natural Sciences of Philadelphia 1936-1937. Part II. - Additional new fishes. Notulae Naturae,(120): 1-7.
Friel, J. 1989. Epidermal keratinization and molting in the banjo catfishes (Siluriformes: Aspredinidae). Abstracts of the 1989 Annual Meeting of the American Society of Ichthyologists and Herpetologists at San Francisco State University, p. 89.
Friel, J. 1992. A phylogenetic revision of Amaralia, a genus of oophagous banjo catfishes. Abstracts of the 1992 Annual Meeting of the American Society of Ichthyologists and Herpetologists at the University of Illinois at Urbana-Champaign, p. 104.
Friel, J. 1994. A phylogenetic study of the Neotropical banjo catfishes (Teleostei: Siluriformes: Aspredinidae) Ph.D.thesis, Duke University, Durham, NC, 256 pp.
Friel, J. 1995. Acanthobunocephalus nicoi, a new genus and species of miniature banjo-catfish from the upper Orinoco and Casiquiare Rivers, Venezuela. (Siluriformes: Aspredinidae). Ichthyological Explorations of Freshwaters, 6(1): 89-95.
Friel, J. & Lundberg, J. G. 1996. Micromyzon akamai, gen. et sp. nov., a small and eyeless banjo catfish (Siluriformes: Aspredinidae) from the river channels of the lower Amazon basin. Copeia, 1996(3): 641-648.
Günther, A. 1864. Catalogue of the Physostomi, containingthe families Siluridae, Characinidae, Haplochitonidae, Sternoptychidae, Scopelidae, Stomiatidae in the collection of the British Museum. Vol. 5, British Museum, London, XXII+ 455 pp.
Howes, G. J. 1983. The cranial muscles of loricarioid catfishes, their homologies and value as taxonomic characters (Teleostei: Siluroidei). Bulletin of the British Museum of Natural History, (Zoology), 45(6): 309-345.
ICZN (International Commission on Zoological Nomenclature). 1985. International code of zoological nomenclature. Third ed. University of California Press, Berkley and Los Angeles, California, 338 pp.
Kner, R. 1855 . Ichthyologische Beiträge. I. Über die Gattungen Aspredo und Chaca C. V. aus der Familie der Welse (Siluroidei). Sitzungsberichte der Mathematisch- Naturwissenschaftlichen Classe der Kaiserlichen Akademie der Wissenschaften, 8: 92-105.
Mees, G. F. 1987. The members of the subfamily Aspredininae, family Aspredinidae in Suriname (Pisces, Nematognathi). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 90(2): 173-192.
Mees, G. F. 1988. The genera of the subfamily Bunocephalinae (Pisces, Nematognathi, Aspredinidae). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 91(1):85-102.
Mees, G. F. 1989. Notes on the genus Dysichthys, subfamily Bunocephalinae, family Aspredinidae (Pisces, Nematognathi). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 92(2): 189-250.
Mo, T. 1991. Anatomy, relationships and systematics of the Bagridae (Teleostei: Siluroidei) with a hypothesis of siluroid phylogeny. Theses Zoologicae 17, Koeltz Scientific Books, Koenigstein, vii + 216 pp., 63 figs.
Myers, G. S. 1960. The genera and ecological geography of the South American banjo catfishes, family Aspredinidae. Stanford Ichthyological Bulletin, 7(4): 132-139.
Pinna, M. C. C. 1993. Higher-level phylogeny of Siluriformes, with a new classification of the order (Teleostei, Ostariophysi). Ph.D. thesis, The City University of New York,xi + 482 pp.
Pinna, M. C. C. 1996. A phylogenetic analysis of the Asiancatfish families Sisoridae, Akysidae and Amblycipitidae, with a hypothesis on the relationships of the neotropical Aspredindae (Teleostei, Ostariophysi). Fieldiana. No. 4:1-83.
Wetzel, J., Wourms, J. and Friel, J. 1977. Modifications of the skin epidermis in the skin-brooding catfish, Platystacus cotylephorus. Environmental Biology of Fishes 50(1):13-25.

About This Page
John P. Friel
Cornell University Museum of Vertebrates,
159 Sapsucker Woods Road,
Ithaca, NY 14850 USA
Page copyright © 1995 John P. Friel
- First online 05 July 1995
Citing this page:
Friel, John P. 1995. Aspredinidae. Banjo Catfishes. Version 05 July 1995 (under construction). http://tolweb.org/Aspredinidae/15208/1995.07.05 in The Tree of Life Web Project, http://tolweb.org/
