



Diapsida
Lizards, Sphenodon, crocodylians, birds, and their extinct relatives
Michel Laurin and Jacques A. Gauthier




This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxPlease note that some authors place the turtles within the Diapsida. Please refer to the accessory page on Diapsid Phylogeny and to the Discussion of Phylogenetic Relationships on the Amniota page for more information on this issue.
Introduction
Diapsids are by far the most speciose group of amniotes, with about 14 600 extant species (Goin et al., 1978). They have invaded all major habitats, from the polar circles (many migratory birds) to deserts (many lizards) and even the ocean (sea snakes, sauropterygians). Diapsids include most flying vertebrates (birds) and most poisonous chordates (snakes and the Gila Monster).
The early history of diapsids is poorly documented. Until the late seventies, the oldest known diapsids were the Upper Permian (250 Myr old) younginiforms from South Africa and Madagascar (Harris and Carroll, 1977; Currie, 1980, 1981, and 1982), and a few other contemporaneous diapsids of uncertain affinities (Carroll, 1976a and b). However, recent work (Reisz, 1977) has extended the fossil record of diapsids to the Pennsylvanian (about 300 Myr ago), and greatly increased our knowledge of the diversity of early diapsids (Brinkman et al., 1984; Reisz et al., 1984; Laurin, 1991; deBraga and Reisz, 1995). The oldest known crown-diapsids (saurians) date from the Late Upper Permian (Carroll, 1975; Evans, 1987).

Figure 1. The lepidosaur Crotaphytus, a terrestrial iguanid from western and central North America. Photograph copyright © 2000 John Merck.
Extant diapsids are classified into either lepidosaurs (lizards and Sphenodon) or archosaurs (birds and crocodiles). Both of these clades are very successful and speciose (Fig. 1), and archosaurs include some of the most fascinating vertebrates that ever lived, such as the pterosaurs (flying reptiles of the Mesozoic) and the many extinct groups of dinosaurs. Indeed, the present diversity of archosaurs, even though it compares favourably with many other clades, is a mere shadow of what it was in the mesozoic.
Diapsida was named after the two fenestrae (holes) found in the temporal region of the skull of most early and some extant diapsids. The lower temporal fenestra is between the jugal, postorbital, squamosal, and quadratojugal. The upper temporal fenestra is between the postorbital, parietal, and squamosal. Some diapsids have lost the lower fenestra (lizards) or even both fenestrae (snakes, amphisbaenids), but their early ancestors had both fenestrae.
Characteristics
Only skeletal autapomorphies (unique derived characters) can be confidently assigned to Diapsida because all the basal lineages of diapsids are extinct. In addition to the presence of an upper and a lower lateral temporal fenestra, these include:
- A suborbital fenestra (Fig. 2). This is a relatively large hole in the palate that is located between the palatine, the ectopterygoid, and the maxilla. Other taxa may have a foramen (a small hole just large enough to allow passage for a nerve or a blood vessel) in this region, but there is usually no fenestra.
- A long radius, measuring between 70% and 90% of the length of the humerus. This ratio is variable in extant diapsids, but early diapsids consistently had a longer radius than other taxa, in which it was typically less than 70% of the humeral length.
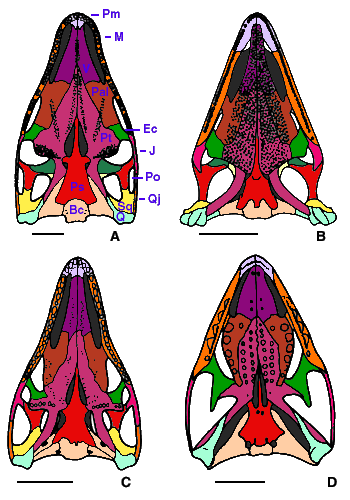
Figure 2. Diapsid skulls in palatal view. A, Petrolacosaurus, a Pennsylvanian araeoscelidian; B, Claudiosaurus, an Upper Permian neodiapsid; C, Youngina, an Upper Permian younginiform; D, Clevosaurus, a Late Triassic sphenodontid (a saurian).
Redrawn from A, Reisz, 1981; B and C, Carroll, 1981; D, Fraser, 1988. Scale bar equals 1 cm. Copyright © 2000 Michel Laurin.
Skeletal autapomorphies can unambiguously be attributed to Sauria, and other characters (soft anatomical, physiological, etc.) present in saurians but not in other extant amniotes are also usually attributed to Sauria, although they may have been present in other diapsids and even in their close relatives ("Protorothyridids" and captorhinids). Saurian autapomorphies include:
- A low concentration of urea in the blood plasma resulting from a loss or suppression of the urea cycle. Lissamphibians, mammals, and turtles have a higher concentration of urea in their plasma.
- The loss of the apposition between the kidney and the adrenal gland. These two organs are juxtaposed in lissamphibians, mammals, and turtles.
- The presence of Huxley's foramen. This foramen is a hole in the distal end of the extracolumella (a cartilaginous extension of the stapes that contacts the tympanum). Huxley's foramen is absent in lissamphibians, mammals, and turtles.
- Temporal muscles originating on the dorsolateral surface of the skull table. In araeoscelidians and younginiforms, the temporal muscles originate from a fascia attached to the lateral edge of the skull table and from the ventral surface of the skull table.
- Prefrontal-nasal suture anterolaterally oriented (Fig. 3D). This suture is parasagittal in araeoscelidians, Claudiosaurus, and younginiforms (Fig. 3A-C).
- Squamosal confined to the dorsal half of the skull, except for a narrow ventral process supporting the quadrate. The squamosal of younginiforms, Apsisaurus, and araeoscelidians is broad ventrally.
- Strong, broad contact between the paroccipital process and the cheek. This contact is weak and often cartilaginous in younginiforms and araeoscelidians.
- Quadrate deeply emarginated posteriorly. The quadrate of saurians supports a tympanum in its deep posterior emargination. The quadrate of younginiforms has a very shallow emargination that probably did not support a tympanum. The quadrate of Apsisaurus and araeoscelidians is not emarginated.
- Slender stapes. The stapes of saurians is modified to function as a middle ear ossicle to transmit high-frequency air-borne sounds from the tympanum to the inner ear. The stapes of younginiforms and araeoscelidians is more massive and it is not specialized as a middle ear ossicle.
- Dorsal process of stapes with ossified connection to paroccipital process of opisthotic. In other taxa, this connection is cartilaginous (or it may consist of a tendon), when it is present.
- Large retroarticular process. This is the insertion point for the muscles that open the lower jaw. The retroarticular process of araeoscelidians, Coelurosauravus, and younginiforms is much smaller.
- Cleithrum absent. The cleithrum is a dermal bone located on the anterior edge of the scapula, dorsal to the clavicle. It is present in araeoscelidians, Coelurosauravus, and younginiforms.
- Lateral manual centrale (a bone in the wrist) small or absent. The lateral and medial centralia are approximately of equal size in araeoscelidians and in some younginiforms primitively (in Acerosodontosaurus).
- Fifth distal tarsal absent (this is a small bone in the ankle, proximal to the fifth toe). Araeoscelidians, Coelurosauravus, and youngina have five distal tarsals.
- Fifth metatarsal hooked (this bone supports the fifth toe). This bone is straight in other diapsids.
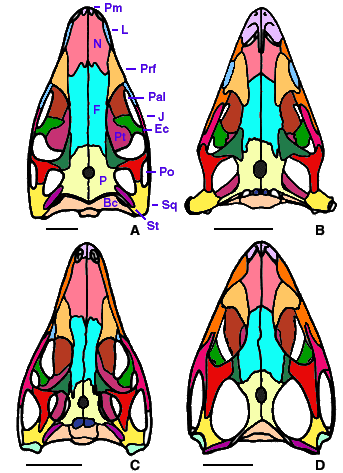
Figure 3. Diapsid skulls in dorsal view. A, Petrolacosaurus, a Pennsylvanian araeoscelidian; B, Claudiosaurus, an Upper Permian neodiapsid; C, Youngina, an Upper Permian younginiform; D, Clevosaurus, a Late Triassic sphenodontid (a saurian).
Redrawn from A, Reisz, 1981; B and C, Carroll, 1981; D, Fraser, 1988. Scale bar equals 1 cm. Copyright © 2000 Michel Laurin.
The linked page Autapomorphies of diapsid clades provides a list of autapomorphies of Neodiapsida and other clades smaller than Diapsida but more inclusive than Sauria.
Discussion of Phylogenetic Relationships
Diapsid phylogeny has been intensively studied in the last decade, but the affinities of many groups are still controversial. For instance, the affinities of the Upper Permian diapsids Galesphyrus (Carroll, 1976a), Heleosaurus (Carroll, 1976b), and Heleosuchus (Carroll, 1987) from South Africa are uncertain because these taxa are represented by fragmentary remains. However, the relationships of the other major groups of stem-diapsids are now reasonably well understood. The affinities of araeoscelidians were initially debated (Peabody, 1952; Vaughn, 1955; Reisz, 1977), but they are now universally believed to be the earliest known group of diapsids (Laurin, 1991; deBraga and Reisz, 1995). Younginiforms were previously believed to represent early lepidosauromorphs (Benton, 1985; Evans, 1988; Gauthier et al., 1988a), but they are now considered to be close relatives of Sauria (Laurin, 1991; Gauthier, 1994).
Some diapsid taxa like Ichthyosauria, Sauropterygia, Placodontia, and Choristodera remain problematic. Placodontia and Choristodera do not appear on the main tree because recent work suggests that they are either lepidosauromorphs or archosauromorphs (Rieppel, 1993, 1994; Rieppel and deBraga, 1996; deBraga and Rieppel, 1997; Merck, 1997). Only ichthyosaurs and sauropterygians appear because they may be either stem-diapsids, archosauromorphs, or lepidosauromorphs (Caldwell, 1996; Mercki, 1997; Motani et al., 1998). Turtles may also be saurians, either lepidosauromorphs or archosauromorphs, but for a discussion of this topic, please refer to the page on Amniota. For more information, see the linked page on Diapsid phylogeny.
Classification of diapsids
Despite numerous studies on diapsid phylogeny and classification, diapsids taxonomy still suffers from a lack of consensus. However, according to the principles of priority suggested by de Queiroz and Gauthier (1990, 1992, 1994), several diapsid taxa have been formally defined phylogenetically, and these definitions should be respected if diapsid taxonomy is ever to be standardized. These taxa include:
- Diapsida: The most recent common ancestor of araeoscelidians, lepidosaurs, and archosaurs, and all its descendants (Laurin, 1991).
- Araeoscelidia: The most recent common ancestor of Araeoscelis and Petrolacosaurus, and all its descendants (Laurin, 1991).
- Neodiapsida: Sauria plus all other diapsids that are closer to saurians than they are to araeoscelidians (Gauthier et al., 1988b).
- Eosuchia: The most recent common ancestor of Coelurosauravus, Apsisaurus, younginiforms, lepidosaurs, and archosaurs and all its descendants (Laurin, 1991).
- Younginiformes: The most recent common ancestor of Youngina, Acerosodontosaurus, and Hovasaurus and all its descendants (Laurin, 1991).
- Sauria: All the descendants of the most recent common ancestor of birds, crocodiles, squamates, and Sphenodon (Gauthier, 1984; Gauthier et al., 1988b).
- Lepidosauromorpha: Extant lepidosaurs and all extinct saurians that are closer to them than they are to extant archosaurs (Gauthier, 1984; Gauthier et al., 1988a).
- Archosauromorpha: Extant archosaurs and all extinct saurians that are closer to them than they are to extant lepidosaurs (Gauthier, 1984, 1994).
Some of these taxa have been given multiple definitions, and some of these definitions have been applied to more than one taxon, but the definitions of taxa given above appear to have priority (de Queiroz and Gauthier, 1990, 1992, and 1994).
This classification, as well as most recent classifications of diapsids, does not recognize Lacertilia as a formal taxon (Lacertilia included lizards but not snakes and amphisbaenids) because this group is not monophyletic (it does not include all the descendants of a common ancestor). Indeed, snakes (Estes et al., 1988; Rieppel, 1988) and amphisbaenids (Wu et al., 1993) are the direct descendants of some lizards, so in this page, the word "lizards" includes these taxa as well.
Hearing in diapsids
The earliest diapsids (and indeed, all known stem diapsids) lacked a tympanum and could not hear high-frequency air-borne sounds, as shown by the lack of a quadrate emargination (the structure that supports the tympanum in most extant diapsids) and the robust shape of their stapes. This indicates that the tympanum present in most saurians developed independently from the tympanum of turtles and mammals (except if turtles are saurians, as recently suggested by Rieppel and deBraga (1996).
A few saurians have lost their tympanum and their ability to hear high-frequency air-borne sounds. These include snakes and many other lepidosaurs, such as Sphenodon, amphisbaenids, Tympanocryptis, Aphaniotes (Barry, 1963). These saurians can only hear low-frequency seismic vibrations (sounds transmitted through the ground).
Early phylogenies of diapsids suggested that the tympanum and the ability to hear high-frequency air-borne sounds had appeared separately in lepidosauromorphs and in archosauromorphs because younginiforms, formerly believed to be the oldest known lepidosauromorphs, apparently lacked a tympanum (Benton, 1985; Evans, 1988). However, the reinterpretation of younginiforms as stem diapsids implies that the tympanum appeared only once in diapsids, probably soon before the divergence between lepidosauromorphs and archosauromorphs (Laurin, 1991).
Diapsid fossil record
Diapsids are known from surprisingly few Paleozoic taxa. Of the few that had been described in the last decades, some have been shown to represent varanopid synapsids, which share several similarities, especially in the postcranial skeleton, but even a few in the skull (such as the presence of a lower temporal fenestra, although this is found in all synapsids), with diapsids. Thus, Apsisaurus, initially described by Laurin (1991) as an Early Permian eosuchian, was recently shown (Reisz et al., 2010) to be a varanopid. Similarly, the Middle Permian form Heleosaurus scholtzi has been shown to be a varanopid (Reisz and Modesto, 2007; Botha-Brink and Modesto, 2009). This leaves only a few taxa, especially the araeoscelidians, younginiforms and Coelurosauravus, in the Permian diapsid fossil record of diapsids.References
Barry T. H. 1963. On the variable occurrence of the tympanum in recent and fossil tetrapods. South African Journal of Science 59: 160-175.
Benton M. J. 1985. Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society 84: 97-164.
Botha-Brink J. & S. P. Modesto. 2009. Anatomy and relationships of the Middle Permian Varanopid Heleosaurus scholtzi based on a social aggregation from the Karoo Basin of South Africa. Journal of Vertebrate Paleontology 29: 389400.
Brinkman D. B., D. S. Berman, & D. A. Eberth. 1984. A new araeoscelid reptile, Zarcasaurus tanyderus, from the Cutler Formation (Lower Permian) of north central New Mexico. New Mexico Geology 1984: 34-39.
Caldwell M. 1996. Ichthyosauria: a preliminary phylogenetic analysis of diapsid affinities. Neuen Jahrbuch für Geologie und Paläontologie. Abhandlungen 200: 361-386.
Carroll R. L. 1975. Permo-Triassic "lizards" from the Karroo. Palaeontologia Africana 18: 71-87.
Carroll R. L. 1976a. Galesphyrus capensis, a younginid eosuchian from the Cistephalus zone of South Africa. Annals of the South African Museum 72: 59-68.
Carroll R. L. 1976b. Eosuchians and the Origin of Archosaurs. In: C. S. Churcher (ed.) Athlon-Essays on Palaeontology in Honour of Loris Shano Russell: 58-79. Toronto: Royal Ontario Museum. Life Sciences. Miscalleneous Publications.
Carroll R. L. 1981. Plesiosaur ancestors from the Upper Permian of Madagascar. Philosophical Transactions of the Royal Society 293: 315-383.
Carroll R. L. 1987. Heleosuchus: an enigmatic diapsid reptile from the Late Permian or Early Triassic of southern Africa. Canadian Journal of Earth Sciences 24: 664-667.
Currie P. J. c. 1980. A new younginid (Reptilia: Eosuchia) from the Upper Permian of Madagascar. Canadian Journal of Earth Sciences 17: 500-511.
Currie P. J. 1981. Hovasaurus boules, an aquatic eosuchian from the Upper Permian of Madagascar. Palaeontologia Africana 24: 99-168.
Currie P. J. 1982. The osteology and relationships of Tangasaurus mennelli Haughton (Reptilia, Eosuchia). Annals of the South African Museum 86: 247-265.
de Queiroz K. & J. Gauthier. 1990. Phylogeny as a central principle in taxonomy: Phylogenetic definitions of taxon names. Systematic Zoology 39: 307-322.
de Queiroz K. & J. Gauthier. 1992. Phylogenetic taxonomy. Annual Review of Ecology and Systematics 23: 449-480.
de Queiroz K. & J. Gauthier. 1994. Toward a phylogenetic system of biological nomenclature. Trends in Ecology and Evolution 9: 27-31.
deBraga M. & R. R. Reisz. 1995. A new diapsid reptile from the uppermost Carboniferous (Stephanian) of Kansas. Palaeontology 38: 199-212.
deBraga M. and O. Rieppel. 1997. Reptile phylogeny and the interrelationships of turtles. Zoological Journal of the Linnean Society 120: 281-354.
Estes R., K. de Queiroz, & J. Gauthier. 1988. Phylogenetic Relationships within Squamata. In: R. Estes and G. Pregill (ed.) Phylogenetic Relationships of the Lizard Families: 119-281. Stanford: Stanford University Press.
Evans S. E. 1987. The phylogeny of Prolacertiformes. In: P. M. Currie and E. H. Koster, Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers in Drumheller, Alberta,
Evans S. E. 1988. The early history and relationships of the Diapsida. In: M. J. Benton (ed.) The phylogeny and classification of the tetrapods: 221-260. Oxford: Clarendon Press.
Fraser N. C. 1988. The osteology and relationships of Clevosaurus (Reptilia: Sphenodontida). Philosophical Transactions of the Royal Society B321: 125-178.
Gauthier J. 1984. A cladistic analysis of the higher systematic categories of the Diapsida. PhD, University of California at Berkeley.
Gauthier J. A. 1994. The diversification of the amniotes. In: D. R. Prothero and R. M. Schoch (ed.) Major Features of Vertebrate Evolution: 129-159. Knoxville, Tennessee: The Paleontological Society.
Gauthier J., R. Estes, & K. de Queiroz. 1988a. A phylogenetic analysis of Lepidosauromorpha. In: R. Estes and G. Pregill (ed.) Phylogenetic relationships of the lizard families: 15-98. Stanford: Stanford University Press.
Gauthier J., A. G. Kluge, & T. Rowe. 1988b. The early evolution of the Amniota. In: M. J. Benton (ed.) The phylogeny and classification of the tetrapods, Volume 1: amphibians, reptiles, birds: 103-155. Oxford: Clarendon Press.
Goin C. J., O. B. Goin, & G. R. Zug. 1978. Introduction to herpetology. Vol. 1. New York: W. H. Freeman and company.
Harriss J. M. & R. L. Carroll. 1977. Kenyasaurus, a new eosuchian reptile from the Early Triassic of Kenya. Journal of Paleontology 51: 139-149.
Laurin M. 1991. The osteology of a Lower Permian eosuchian from Texas and a review of diapsid phylogeny. Zoological Journal of the Linnean Society 101: 59-95.
Merck J. 1997. A phylogenetic analysis of the euryapsid reptiles. Journal of Vertebrate Paleontology 17: 65A.
Motani R., N. Minoura, and T. Ando. 1998. Ichthyosaurian relationships illuminated by new primitive skeletons from Japan. Nature 393: 255-257.
Peabody F. E. 1952. Petrolacosaurus kansensis Lane, a Pennsylvanian reptile from Kansas. Paleontological Contributions. University of Kansas. 1: 1-41.
Reisz R. R. 1977. Petrolacosaurus, the Oldest Known Diapsid Reptile. Science 196: 1091-1093.
Reisz R. R. 1981. A diapsid reptile from the Pennsylvanian of Kansas. University of Kansas Publications of the Museum of Natural History 7: 1-74.
Reisz R. R. & S. P. Modesto. 2007. Heleosaurus scholtzi from the permian of South Africa: a varanopid synapsid not a diapsid reptile. Journal of Vertebrate Paleontology 27: 727-733.
Reisz R. R., D. S. Berman, & D. Scott. 1984. The anatomy and relationships of the Lower Permian reptile Araeoscelis. Journal of Vertebrate Paleontology 4: 57-67.
Reisz R. R., M. Laurin, & D. Marjanović. 2010. Apsisaurus witteri from the Lower Permian of Texas: yet another small varanopid synapsid, not a diapsid. Journal of Vertebrate Paleontology 30: 16281631.
Rieppel O. 1988. The classification of the Squamata. In: M. J. Benton (ed.) The Phylogeny and Classification of the Tetrapods: 261-293. Oxford: Clarendon Press.
Rieppel O. 1993. Euryapsid relationships: A preliminary analysis. Neuen Jahrbuch für Geologie und Paläontologie. Abhandlungen 188: 241-264.
Rieppel O. 1994. Osteology of Simosaurus gaillardoti and the relationships of stem-group sauropterygia. Fieldiana Geology 1462: 1-85.
Rieppel O. & M. deBraga. 1996. Turtles as diapsid reptiles. Nature 384: 453-455.
Vaughn P. P. 1955. The Permian reptile Araeoscelis restudied. Bulletin of the Museum of Comparative Zoology 113: 305-467.
Wu X.-c., D. B. Brinkman, A. P. Russell, Z.-m. Dong, P. J. Currie, L.-h. Hou, & G.-h. Cui. 1993. Oldest known amphisbaenian from the Upper Cretaceous of Chinese Inner Mongolia. Nature 366: 57-59.
Information on the Internet
- Those Diverse Diapsids. UCMP Berkeley.
- Mystery of the Oldest Flying Vertebrate . Information about Coelurosauravus. Royal Ontario Museum.
- Introduction to the Ichthyosauria. UCMP Berkeley.
- Ichthyosaur Page. Ryosuke Motani.
- The Plesiosaur Site. Richard Forrest.
- Plesiosaur FAQ's. Raymond Thaddeus C. Ancog.
- Something about Plesiosaurs. Mike Everhart.
Title Illustrations

| Scientific Name | Spinoaequalis schultzei, Haptodus garnettensis, Sigillaria |
|---|---|
| Comments | The Pennsylvanian diapsid Spinoaequalis schultzei jumps into the water to escape a hungry Haptodus garnettensis (a synapsid). The base of a Sigillaria trunk is visible in the upper right corner. |
| Creator | Michael deBraga |
| Copyright | © 2000 Michael deBraga |
About This Page
Michel Laurin

Muséum National d'Histoire Naturelle, Paris, France
Jacques A. Gauthier
Yale University, New Haven, Connecticut, USA
Correspondence regarding this page should be directed to Michel Laurin at
Page copyright © 2012 Michel Laurin and Jacques A. Gauthier
 Page: Tree of Life
Diapsida. Lizards, Sphenodon, crocodylians, birds, and their extinct relatives.
Authored by
Michel Laurin and Jacques A. Gauthier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Diapsida. Lizards, Sphenodon, crocodylians, birds, and their extinct relatives.
Authored by
Michel Laurin and Jacques A. Gauthier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 22 June 2000
- Content changed 20 April 2011
Citing this page:
Laurin, Michel and Jacques A. Gauthier. 2011. Diapsida. Lizards, Sphenodon, crocodylians, birds, and their extinct relatives. Version 20 April 2011. http://tolweb.org/Diapsida/14866/2011.04.20 in The Tree of Life Web Project, http://tolweb.org/

