




Synapsida
Mammals and their extinct relatives
Michel Laurin and Robert R. Reisz




This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
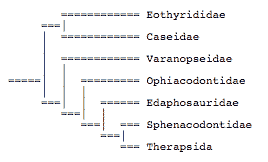
close boxPhylogeny of synapsids, after Reisz (1986) and Laurin (1993). Terminology after Kemp (1982), Reisz (1986), and Reisz et al. (1992).
Introduction
Synapsids include mammals and all extinct amniotes more closely related to mammals than to reptiles. Synapsids are the dominant large terrestrial animals worldwide, and they have also invaded the oceans (whales, pinnipeds) and the air (bats). The oldest known synapsid is an ophiacodontid from the Middle Pennsylvanian (320 million years ago) of Joggins, Nova Scotia. By the Lower Permian, therapsids (the group that includes mammals and most of their Upper Permian and more recent relatives) had appeared (Laurin and Reisz, 1990, 1996; Amson and Laurin, 2011). However, the oldest known mammal only dates back to the Jurassic (Rowe, 1988). The fossil record of synapsids is one of the most extensive of any groups of vertebrates. This fossil record has been used to illustrate the concept of evolution (Hopson, 1987) and to test macroevolutionary patterns (Kemp, 1985). The largest gap in this fossil record is between the Permo-Carboniferous synapsids and therapsids.
The fossil record provides conclusive evidence that synapsids are the first amniotes to diversify. Synapsids quickly became the most diverse, widespread and most common amniotes in the Late Carboniferous, and they maintained this predominant position throughout the Paleozoic. Only during the Early Mesozoic are the synapsids eclipsed by the evolutionary radiation of reptiles (Benton, 1983; Charig, 1984). Within the Late Carboniferous and Early Permian, two different herbivorous and several faunivorous synapsid groups can be recognized.
The fossil record suggests that during the Carboniferous and Early Permian, synapsids and other amniotes were restricted to the paleoequatorial and subequatorial regions. During the Late Permian, the distribution of synapsids, and of some of the other amniotes becomes cosmopolitan. However, the evidence for this pattern rests on rather weak negative evidence (i. e., no Permo-Carboniferous synapsids have been found outside paleoequatorial regions, but other areas have not been prospected intensively).
Early synapsids had a sprawling posture and a small brain, like most early tetrapods. The parasagittal gait characteristic of most mammals (Fig. 1) appeared gradually because some therapsids were apparently capable of sprawling and parasagittal gait, and this character may have appeared in the hind limb before the fore limb. Early synapsids were moderately large (body length between 50 cm and 3 m) and most were carnivorous or insectivorous, but caseids and Edaphosaurus were herbivorous. Mammals include the largest tetrapods that ever lived (the blue whale is larger than any dinosaur), as well as very small species.
Most mammals are viviparous, but the platypus (a monotreme) is oviparous, and most, if not all, non-mammalian synapsids were probably oviparous. All mammals have mammary glands, but the presence of these structures cannot be determined in extinct taxa. The oldest preserved synapsid hair appears to belong to a Middle Jurassic docodontan mammaliaform (a stem-synapsid) from China (Ji et al., 2006). However, hair is infrequently fossilized and the next oldest record is from a Paleocene multituberculate (Meng and Wyss, 1997). Therefore, its real date of appearance is unknown. The position of this group has been debated, but they are probably mammals (Rowe, 1988), so the presence of hair in multituberculates was predictible.

Figure 1. An extant synapsid (a black-tailed deer). Like most of the
other 3500 extant synapsid species, it is endothermic and has fur, but these
features were certainly absent in all Paleozoic synapsids. Photograph by Roger Laurin.
Characteristics
All known early synapsids had a lateral temporal fenestra (a hole in the lateral surface of the skull behind the orbit) that is still present in a modified form in mammals. Primitively, the lateral temporal fenestra was bordered by only three bones (Fig. 2).
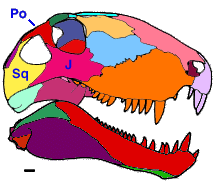
Figure 2. Temporal fenestration primitively found in synapsids.
Abbreviations: J, jugal; Po, postorbital; Sq, squamosal. Scale equals 1 cm.
The occiput was broad and it faced posterodorsally. In other early amniotes, the endochondral bones of the braincase formed a narrower surface, and it was more or less vertical.
The pubis had a long, ridged anterodorsal edge.
Physiology
The earliest synapsids were probably ectothermic (their body temperature depended on the environment), as shown by the presence of a large "sail" on the back of edaphosaurids and some sphenacodontids. This sail was composed of greatly elongated neural spines that was grooved anteriorly and posteriorly to accommodate blood vessels, and skin probably connected all the neural spines together. This sail may have allowed these early synapsids to raise their body temperature faster than similarly-sized tetrapods lacking a sail. It was also thought that the sail was used to radiate excess heat into the environment, but recent studies suggest otherwise (Haack, 1986). The presence of the sail suggests that early synapsids lacked hair. Hair usually does not fossilize, but its presence in most monotremes, marsupials, and placentals suggests that it was present in their last common ancestor. Endothermy is also difficult to determine in extinct taxa, but it probably appeared slightly before the first mammal was born. The presence of epidermal horny scales cannot be determined in most fossil taxa, but bony ventral scales were primitively present, as shown by their presence in most reasonably complete fossils of early synapsids.
Discussion of Phylogenetic Relationships
Early synapsid phylogeny is relatively uncontroversial, except for the therapsid affinities of Tetraceratops (Amson and Laurin, 2011). Recent studies by Reisz (1980, 1986), Brinkman and Eberth (1983), Gauthier et al. (1988), and Laurin (1993) have resolved the relationships between the largest groups of early synapsids. More recent studies by Reisz et al. (1992) and Berman et al. (1995) have confirmed these findings.
Reisz (1980) produced the first phylogeny of synapsids using phylogenetic systematics. He argued that most carnivorous synapsids (ophiacodontids, varanopseids, sphenacodontines, and therapsids formed a clade that excluded the herbivorous edaphosaurids and caseids.
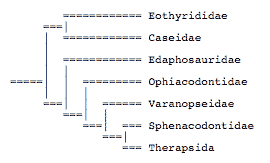
Brinkman and Eberth (1983) argued that varanopseids were closely related to caseids and that edaphosaurids were closely related to sphenacodontids:
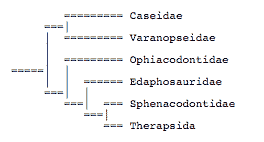
Reisz (1986) agreed with Brinkman and Eberth (1983) that edaphosaurids were closely related to sphenacodontids, but he argued that varanopseids were more closely related to ophiacodontids than to caseids:
This new position of varanopseids has not been directly questioned by subsequent studies. The phylogeny of Gauthier et al. (1988) resembled that of Brinkman and Eberth (1983), but they did not test the monophyly of the varanopseid-caseid clade.
Gauthier et al. (1989), Hopson (1991), and Reisz et al. (1992) suggested that Haptodus, previously believed to be a sphencodontid, did not belong to this group:
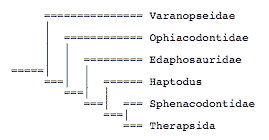
Laurin (1993) studied the relationships of several problematic taxa previously called haptodontines. He suggested that haptodontines were a paraphyletic group:
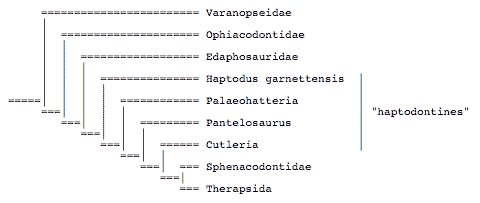
See the page on Evolution of early synapsids for synapomorphies justifying the latest phylogenies and for anatomical drawings.
References
Amson E. & M. Laurin. 2011. On the affinities of Tetraceratops insignis, an Early Permian synapsid. Acta Palaeontologica Polonica 56: 301312.
Benton M. J. 1983. Dinosaur success in the Triassic: a noncompetitive ecological model. Quarterly Review of Biology 58: 29-55.
Berman D. S., R. R. Reisz, J. R. Bolt, and D. Scott. 1995. The cranial anatomy and relationships of the synapsid Varanosaurus (Eupelycosauria: Ophiacodontidae) from the Early Permian of Texas and Oklahoma. Annals of Carnegie Museum 64: 99-133.
Brinkman, D. and D. A. Eberth. 1983. The interrelationships of pelycosaurs. Breviora 473: 1-35.
Charig A. 1984. Competition between therapsids and archosaurs during the Triassic period: a review and synthesis of current theories. In: M. W. J. Ferguson (ed.) The structure, development and evolution of reptiles: 597-628. London: Academic Press.
Gauthier, J., A. G. Kluge, and T. Rowe. 1988. Amniote phylogeny and the importance of fossils. Cladistics 4: 105-209.
Gauthier J., D. C. Cannatella, K. De Queiroz, A. G. Kluge, and T. Rowe. 1989. Tetrapod phylogeny. In: B. Fernholm, K. Bremer, and H. Jornvall (ed.) The Hierarchy of Life: 337-353. New York: Elsevier Science Publishers B. V. (Biomedical Division).
Haack, S. C. 1986. A thermal model of the sailback pelycosaur. Paleobiology 12: 450-458.
Hopson J. A. 1987. The Mammal-like Reptiles. A study of transitional fossils. The American biology teacher 49: 16-26.
Hopson J. A. 1991. Systematics of the nonmammalian Synapsida and implications for patterns of evolution in synapsids. In: H.-P. Schultze and L. Trueb (ed.) Origins of the higher groups of tetrapods—Controversy and Consensus: 635-693. Ithaca: Cornell University Press.
Ji Q., Z.-X. Luo, C.-X. Yuan, & A. R. Tabrum. 2006. A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311: 1123-1127.
Kemp, T. S. 1982. Mammal-like Reptiles and the Origin of Mammals. Academic press, New York, 363 pp.
Kemp T. S. 1985. Synapsid reptiles and the origin of higher taxa. Special Papers in Palaeontology 33: 175-184.
Laurin, M. 1993. Anatomy and relationships of Haptodus garnettensis, a Pennsylvanian synapsid from Kansas. Journal of Vertebrate Paleontology 13: 200-229.
Laurin, M. and R. R. Reisz. 1990. Tetraceratops is the oldest known therapsid. Nature 345: 249-250.
Laurin M. and R. R. Reisz. 1996. The osteology and relationships of Tetraceratops insignis, the oldest known therapsid. Journal of Vertebrate Paleontology 16: 95-102.
Meng J. and A. R. Wyss. 1997. Multituberculate and other mammal hair recovered from Palaeogene excreta. Nature 385: 712-714.
Reisz R. R. 1980. The Pelycosauria: A review of phylogenetic relationships. In: A. L. Panchen (ed.) The Terrestrial Environment and the Origin of Land Vertebrates: 553-592. London: Academic Press.
Reisz, R. R. 1986. Pelycosauria. Encyclopedia of Paleoherpetology 17A: 1-102.
Reisz, R. R., D. S. Berman, and D. Scott. 1992. The cranial anatomy and relationships of Secodontosaurus, an unusual mammal-like reptile (Pelycosauria: Sphenacodontidae) from the early Permian of Texas. Zoological Journal of the Linnean Society 104: 127-184.
Rowe T. 1988. Definition, diagnosis, and origin of Mammalia. Journal of Vertebrate Paleontology 8: 241-264.
Zimmer C. Meet the relatives. Discover, 1991, 56-57.
Information on the Internet
For a general introduction to mammals see the UCMP Berkeley Hall of Mammals and Animal Diversity Web: Class Mammalia
Title Illustrations

| Scientific Name | Tetraceratops |
|---|---|
| Location | Texas |
| Comments | Tetraceratops, the oldest known therapsid. This strange synapsid, found in Texas in the last century, has recently been reinterpreted as the only known Lower Permian therapsid (Laurin and Reisz, 1990). |
| Reference | Zimmer C. 1991. Meet the relatives. Discover, 1991, 56-57 |
| Creator | Drawing by Steve Kirk |
| Specimen Condition | Fossil -- Period: Lower Permian |
| Copyright | © 1991 Discover Magazine |
About This Page
Michel Laurin

Muséum National d'Histoire Naturelle, Paris, France
Robert R. Reisz
University of Toronto, Mississauga, Ontario, Canada
Correspondence regarding this page should be directed to Michel Laurin at
Page copyright © 2012 Michel Laurin and
 Page: Tree of Life
Synapsida. Mammals and their extinct relatives.
Authored by
Michel Laurin and Robert R. Reisz.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Synapsida. Mammals and their extinct relatives.
Authored by
Michel Laurin and Robert R. Reisz.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 07 May 1997
- Content changed 14 August 2011
Citing this page:
Laurin, Michel and Robert R. Reisz. 2011. Synapsida. Mammals and their extinct relatives. Version 14 August 2011. http://tolweb.org/Synapsida/14845/2011.08.14 in The Tree of Life Web Project, http://tolweb.org/

