



Reithrodon
Reithrodontini, Coney rats, Bunny rats
Scott J. Steppan and Michael Reno




This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Reithrodontini is composed of a single genus with two recognized species: R. typicus and R. auritus. Reithrodon typicus is the type species (Waterhouse, 1837). They are commonly known as the coney or bunny rats and are found mostly in southern South America from Tierra del Fuego to northern Argentina (Pardiñas and Galliari, 2001). The Reithrodontini are viewed as agricultural pests because they consume largely only grasses; some have been observed consuming amounts equal to their body weight in a single night (Pearson, 1988).

Reithrodon. © O. Pearson
Characteristics
Reithroidon are large, robust rodents with an average body mass of 80 grams (Pearson, 1988). Fur is usually dense and gray in color. The tail, which is slightly longer than the body, is covered with hair and ends with a tuft (Gyldenstolpe, 1932). The ears are large, rounded, and covered with hair. R. typicus may be differentiated from R. auritus by the sole of its feet with are bare. R. auritus has hair on the soles of its feet (Thomas, 1920).
The Reithrodontini range from Tierra del Fuego to the Pataganoian region of Argentina. They are found in patchy clusters in the northern regions of Argentina (Pardiñas and Galliari, 2001). They prefer the open habitat of grassy prairies but have been found in a great variety of habitats ranging from grassland to woodland areas (Pearson, 1988; Johnson et al., 1990). They are known to form tunnels in tall grass and even cohabitate these tunnels with other rodent, such as Akodon longipilus (Pearson, 1988). The Reithrodontini can be either diurnal or nocturnal depending on the weather conditions; they are active year round (Mann Fischer, 1978). They are reproductively active in the spring with litter sizes ranging from one to eight (Pearson, 1988; Pine et al., 1979).
Morphology
The Reithrodontini are large rodents with large, rounded ears which are covered by hair. They have a tail slightly longer than their body. The hind legs are long and have hair between the sole pads. The first and fifth digits of the foot are shorter than the middle three (Glydenstolpe, 1932; Pearson, 1988, 1995). The incisors have two grooves, and the molars are hypsodont (high crowned) and laminated. The cusps and crests of the third molar are arranged in a sigmoidal array (S-haped), and the mesoloph is absent (Pearson, 1995; Steppan, 1995). The Reithrodontini can easily be distgiushed from other Sigmodontinae by regular and rhomboidal cuticular scales on dorsal guard hairs (Busch, 1986).

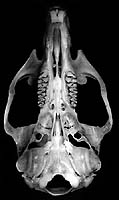

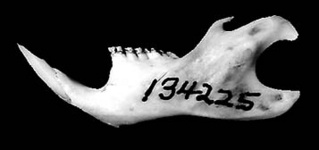
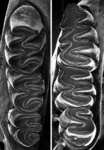
Reithrodon auritus. © Scott J. Steppan
Compare the skulls, jaws, or molars of all phyllotine genera by selecting the appropriate view below.
| Dorsal | Ventral | Lateral | Jaw | Teeth |
There are two characteristics which differentiate R. typicus and R. auritus: karyotyping and hair on the sole pads. R. typicus has a diploid chromosome number of 28 and no hair on the sole pads while R. auritus has a diploid chromosome number of 34 and hair on its sole pads (Thomas,1920; Freitas et al., 1983).
Taxonomy
Waterhouse (1837) first described the genus Reithrodon. Several taxa, such as Euneomys and Irenomys, were initally thought to be members of Reithrodon (Waterhouse, 1839; Trouessart, 1898), but were later removed to their own genera (Thomas, 1901; Allen 1903). For many years R. cuniculoides was recognized as the type species (Coues 1874; Trouessart, 1905); however in 1926, a change in International Rules of Nomenclature placed R. typicus as type species.
Herskovitz (1955) placed the Reithrodontini within a sigmodont group along with Sigmodon, Holochilus, Sigmodon, and Neotomys. Karyotyping showed the Reithrodontini to be more closely related to the Phyllotini (Pearson and Patton, 1976). Olds and Anderson (1989) placed the Reithrodontini in an informal reithrodont group along with Euneomys and Neotomys within the Phyllotini. This would be further supported by other studies (Steppan 1993; Steppan and Pardiñas, 1998; Ortiz et al., 2000).
Recent phylogenetic studies have removed Reithrodon from the Phyllotini and elevated them first to a suprageneric group (Engel et al., 1998) and later to tribe status (Smith and Patton, 1999). D’Elía (2003) further confirmed this through the use of interphotoreceptor retinoid binding protein (IRBP), a nuclear gene. More recent studies using multiple nuclear genes have been in agreement (Steppan, unpublished).
References
Allen, J. A. 1903. Descriptions of new rodents from southern Patagonia, with a note on the genus Euneomys Coues, and an addendum to Article IV, on Siberian mammals. Bulletin of the American Museum of National History 19:185-195.
Coues, E. 1874. Synposis of the Muridae of North America. Proceedings of the Academy of Natural Sciences, Philadelphia 26:173-196.Hershkovitz, P. 1955. South American march rats, genus Holochilus, with a summary of sigmodont rodents. Field Museum of Natural History, Zoology Series 27:639-687.
DElia, G. 2003. Phylogenetics of Sigmodontinae (Rodentia, Muroidea, Cricetidae), with special references to the akodont group, and with additional comments on historical biogeography. Cladistics 19:307-323.
Engel, S. R., K. M. Hogan, J. F. Taylor, and S. K. Davis. 1998. Molecular systematics and paleogeography of the South American sigmodontine rodents. Molecular Biology and Evolution 15:35-49.
Freitas, P. R. O., M. Mattevi, and L. F. B. Oliveira. 1983. G- and C-band karyotype of Reithrodon auritus from Brazil. Journal of Mammalogy 64:318-321.
Johnson, W. E., W. L. Franklin, and J. A. Iriarte. 1990. The mammalian fauna of the northern Chilean Patagonia: a biogeographical dilemma. Mammalia 54:457-469.
Ortiz, P. E., U. F. J. Pardiñas, and J. E. Powell. 1998. Roedores del Pleistoceno-Holoceno del Valle de Tafi (Tucuman, Argentina). Aspectos sistimaticos y paleoambientales. Acta Geologica Lilloana, Tucuman 18:173-174.
Pardiñas, U. F. J. and C. A. Galliari. 2001. Reithrodon auritus. Mammalian Species 664:1-8.
Pearson, O. P. 1988. Biology and feeding dynamics of a South American herbivorous rodent, Reithrodon. Studies on Neotropical Fauna and Environment 23:25-39.Gyldenstolpe, N. 1932. A manual of Neotropical sigmodont rodents. Kunglia Svenska Vetenskapsakademiens Handlingar, Stockholm, Serie 3, 11(3):1-164.
Pearson, O. P. 1995. Annotated keys for identifying small mammals living in or near Nahuel Huapi National Park or Lanin National Park, southern Argentina. Mastozoologia Neotropical 2:99-148.
Pearson, O. P. and J. L. Patton. 1976. Relationships among South American phyllotine rodents based on chromosomal analysis. Journal of Mammalogy 57:339-350.
Pine, R. H., S. D. Miller, and M. L. Schamberger. 1979. Contributions to the mammalogy of Chile. Mammalia 43:339-376.
Smith, M. F. and J. L. Patton. 1999. Phylogenetic relationships and the radiation of sigmodontine rodents in South America: evidence from cytochrome b. Journal of Mammalian Evolution 6:89-128.Olds, N. and S. Anderson. 1989. A diagnosis of the tribe Phyllotini (Rodentia: Muridae) Pp 55-75 in Advances in Neotropical mammalogy (K. H. Redford and J. F. Eisenberg, eds.). Sanhill Crane Press, Gainesville, Florida.
Steppan, S. J. 1993. Phylogenetic relationships among the Phyllotini (Rodentia: Sigmodontinae) using morphological characteristics. Journal of Mammalian Evolution 1:187-213.
Steppan, S. J. 1995. Revision of the tribe Phyllotini (Rodentia: Sigmodontinae), with a phylogenetic hypothesis for the Sigmodontinae. Fieldiana: Zoology, New Series 80:1-112.Busch, M. 1986. Identificacion de algunas especices de pequeños mamiferos de la provincial de Buenos Aires mediante caracteristicas de sus pelos. Physis 44(107):113-118.
Steppan, S. J. and Pardiñas. 1998. Two new fossil muroids (Sigmodontinae: Phyllotini) from the Early Pleistocene of Argentina: phylogeny and paleoecology. Journal of Vertebrate Paleontology 18:640-649.
Thamas, O. 1920. New species of reithrodon, Abrocoma, and Scapteromys from Argentina. Annals and Magazine of Natural History, Series 9, 5:473-478.
Thomas, O. 1901. Ann. Mag. Nat. Hist., (7) VIII, p. 254.
Trouessart, E. 1898. Cat. Mamm. Viv. Foss., Supplement.
Trouessart, E. 1905. Cat. Mamm. Viv. Foss., Supplement.
Waterhouse, G. R. 1837. [Characters of new species of the genus Mus, from the collection of Mr. Darwin.] Proceedings of the Zoological Society of London 1837:15-21, 27-32.
Waterhouse, G. R. 1839. Zoology Voyage Beagle. P. 68.
Information on the Internet
Mammal Species of the World: a Taxonomic and Geographic Reference. Internet version.Title Illustrations

| Scientific Name | Reithrodon auritus |
|---|---|
| Copyright | © 1997 O. Pearson |
About This Page
Scott J. Steppan

Florida State University, Tallahassee, Florida, USA
Michael Reno
Florida State University, Tallahassee, Florida, USA
Correspondence regarding this page should be directed to Scott J. Steppan at
Page copyright © 2007 Scott J. Steppan
All Rights Reserved.
- First online 01 October 1996
- Content changed 08 January 2007
Citing this page:
Steppan, Scott J. and Michael Reno. 2007. Reithrodon. Reithrodontini, Coney rats, Bunny rats. Version 08 January 2007 (under construction). http://tolweb.org/Reithrodon/16700/2007.01.08 in The Tree of Life Web Project, http://tolweb.org/

