Origin of the filaments
The primitive (plesiomorphic) arrangement of arms in coleoid cephalopods is thought to be 10 equal arms. This arrangement is known in some fossil coleoids (belemnoids) and the presence of ten unequal arms in modern decapods is easily derived from such a condition. Therefore, octopods, with eight arms, have apparently lost one pair. If the vampire filaments represent modified arms II and if this is the pair that is lacking in octopods, then strong support would exist for a vampire-octopod affinity. Embryological evidence suggests that the missing arm pair in octopods is either arms II or III (Boletzky, 1978-79).
Pickford (e.g., 1940) concluded that the filaments were modified arms II based on their position in series with the arms, venous connections and innervation. Young (1967) clarified the innervation and argued that the filaments were not arms but homologues of the preocular tentacles of Nautilus. The axial nerves of arms send most of their fibers to the anterior subesophageal mass (brachial lobe) of the brain. Young found that the filament nerves bypass this lobe with efferent fibers arising from the middle subesophageal mass and the afferent fibers reaching an enlarged ventral magnocellular lobe. Dilly, et al. (1977), however, demonstated that large nerve bundles from the tentacles of the deep-sea squid Mastigoteuthis also pass to an enlarged ventral magnocellular lobe. Presumably the combined sensory function of thousands of suckers carried by the tentacles of Mastigoteuthis requires the large nerve bundle and the enlarged ventral magnocellular lobe. Similarly the still poorly understood functions of the filaments of Vampyroteuthis probably emphasized the connection to and size of the ventral magnocellular lobe while a loss of coordination with the principal arms resulted in a great reduction (at least) in the connection to the anterior subesophageal mass. As a result, the innervation of the filaments became a weak argument against filament-arm homology.
The homology of the filament appeared to be resolved when J. Z. Young (1977) reported a connection between the axial nerve of the vampire filament and ganglia of the circumoral commissure that connects the axial nerves of all arms. This evidence, however, could not be confirmed by Young and Vecchione (1996).
Recent confirmation of Pickford's earlier evidence (Pickford, 1946) that the vampire hatchling has relatively thick (i.e., more arm-like) filaments supports the homology between arms and filaments (Young and Vecchione, 1999).

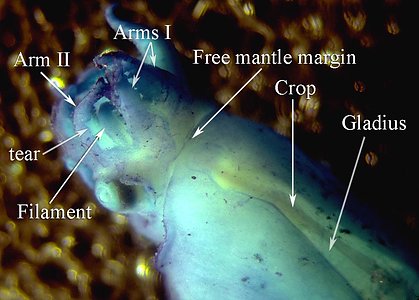
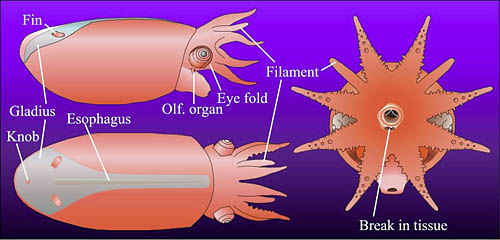
Figure. Vampyroteuthis infernalis hatchling. Figures modified from Young and Vecchione, 1999. Top - Dorsal and slightly lateral photograph of the lightly stained (methylene blue) hatchling. Bottom - Lateral, dorsal and anterior drawings of the same hatchling. Morphologically the specimen is an advanced embryo with a large internal yolk sac.
The weight of evidence indicates that the filaments represent arms II. Proof may depend on examining the embryonic development of the vampire squid which is not feasible at present.
The filament
Each long filament has muscles that run the along one side. Contraction of these muscles causes coiling of the filament and its retraction into the small filament pocket that lies at the base of the web. The following images show a filament that has been removed from a recently dead vampire. Most of the filament has an orange pigmentation; the cut base (arrow), however, is abruptly broader and has a pink color. Much of the tight coiling has been disrupted by handling.
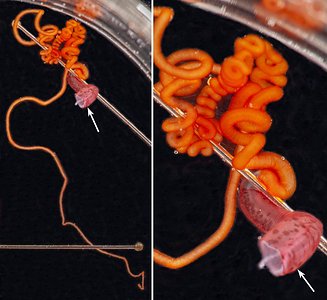
Figure. Filament of Vampyroteuthis infernalis. Left - The entire filament from the cut base (arrow) to the filament tip. Right - Enlargement of the upper part of the left image showing how tight some of the undisturbed coils can be. Photographed in a small dish aboard ship by Henk-Jan Hoving.
Under the dissecting microscope numerous, straight "hairs" are seen extending from the filament. These "hairs" are thought to aid in capturing food.
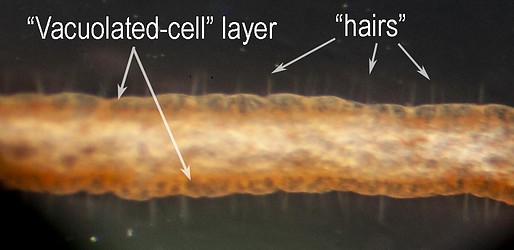
Figure. V. infernalis filament. Photomicrograph of a portion of the intact, fresh filament. Photograph by H. Hoving.
The Histology of the filaments has been examined by Hoving and Robison, 2012. The "hairs" could not be detected in the paraffin-embedded sections but could be seen in SEM photographs collapsed onto the surface of the filament; there, they appear to be tree-like branching structures rather than simple hairs. The surface of the filament consists of a layer of "vacuolated" cells with little internal structure. In SEM photographs, these cells are also collapsed and the "hairs" seem to arise at the junctures between these cells. Beneath the vacuolated cell layer is a layer of simple cuboidal cells with larger cells broadly interspersed. These larger cells are apparently nerve cells as they have extensions that pass into a large, axial nerve. A large muscle exists along one side of the filament and its contraction results in the coiling of the filament.
The "hairs" apparently secure food particles (Hoving and Robison, 2012). The retraction of the filaments is a slow process and the vampire squid may swim to the portion of the tentacle holding the food (Hunt, 1996) and enclose it within the web. Next, the filament is drawn between the arms, wrapped in mucous produced by cells on the suckers, sucker stalks and cirri, and passed to the mouth (Hoving and Robison, 2012).




