The amoeboid body form is one of the most widespread. Not only does it occur in amoebae, but it is also found in algae, fungi and animals. From comparisons of molecular data and from life history data, it is clear that the amoeboid body form is an adaptive form and that the amoebae are polyphyletic.
Traditionally, the amoebae have been classified as the protozoan taxon Sarcodina, with a taxonomic structure that separated species with supported pseudodpodia from those with unsupported pseudopodia, and those with skeletal elements, from those without. Older schemes such as this grouped together related taxa and exluded other related taxa. The amoebae are probably the last large area of protists which have to be fully documented and explored, although progress in that area is now taking place (Amaral, Silberman).
The "Sarcodina" included the following taxa:
- Actinopoda (with stiff pseudopodia)
- Radiolaria (radiolaria)
- Acantharea (acantharea)
- Polycystinea (polycystines)
- Phaeodarea (phaeodarea)
- Heliozoa (heliozoa)
- Actinophryida (actinophryids)
- Centohelida (centrohelids)
- Desmothoracida (desmothoracids)
- Gymnosphaerida (gymnosphaerids)
- Taxopodida (Sticholonche)
- Radiolaria (radiolaria)
- Rhizopoda (with unsupported pseudopodia)
- Lobosea (lobose amoebae)
- Gymnamoebae (naked lobose amoebae)
- Testacealobosa (testate lobose amoebae)
- Filosea (filose amoebae)
- Aconchulinida (naked filose amoebae)
- Testaceafilosea (testate efilose amoebae) )
- Granuloreticulosea (amoeboae with pseudopodia forming a network)
- Foraminiferida (foraminifera)
- Lobosea (lobose amoebae)
The amoebae are of importance both medically and ecologically. Naegleria is the cause of amoebic meningitis; other species are more or less benign coresidents in digestive systems of ourselves or other animals or can facultatively attack the conjunctiva of our eyes. In marine ecosystems, radiolaria and foraminifera are significant members of communities, both as consumers and as producers (many planktonic marine amoebae and foraminifera from intertidal habitats harbour large numbers of symbiotic algae). Naked amoebae are major consumers of bacteria in soil ecosystems and are believed to occupy the same key role there as flagellates do in aquatic ecosystems (that of recycling bacterial productivity and ensuring nutrient regeneration and continued functioning of the ecosystem).

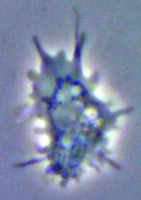
Mayorella (Gymnamoebae) a naked amoeba with conical pseudopodia, living cell.
Amoebae are frequently maligned as shapeless blobs. Despite their evident pleomorphism which comes from being able to emit and resorb extensions of cytoplasm, active amoebae do have a front and a back.
Pseudopodia. Different categories of amoebae can be distinguished by the appearance of the pseudopodia, and these are summarised in the diagram below. Pseudopodia may have internal microtubules which give them rigidity (as in radiolaria, heliozoa and foraminifera). The stiffened pseudopodia of radiolaria and heliozoa tend to extend radially and are called actinopoda. Those of foraminifera branch and fuse, and because of the resulting net-like structure, are referred to as granulo-reticulose pseudopodia. Unsupported pseudopodia may be lobose (broad) or filose (thread-like), many may be produced at the same time (polypodial) or cells may migrate with a single advancing pseudopodium (monopodial).
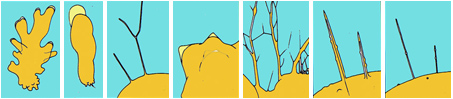
The forms of pseudopodia: from left: polypodial and lobose, monopodial and lobose, filose, conical, granuloreticulose, tapering actinopods, non-tapering actinopods.
The posterior end of lobose amoebae may also be distinctive, and is referred to as the uroid. The form of this region may be distinctive.
Ultrastructure. As with other protists, details of nuclear division profiles, mitochondria, extrusomes and other inclusions have helped to establish the identities for categories of amoebae - especially the heliozoa and radiolaria. The ramicristates are a major subset of amoeboid organisms distinguished by having branched cristae within the mitochondria.
Life cycle: amoebae typically grow and divide; this pattern of vegetative growth may or may not include encysted states, flagellated forms, or a transition from a sexual to an asexual phase of growth.
The amoeboid body form is widespread among the protists. The following groups are entirely or predominantly amoeboid - except where specified.
Groups of Heterotrophic Amoebae
| Name of group | Organization | Composition / sample genera |
|---|---|---|
| Acantharea | radiolaria | Very diverse, e.g. Acanthometron |
| Biomyxa | Named amoebae with thin anastomosing pseuodopodia | One genus |
| Centroheliozoa | heliozoa | About 10 genera, e.g. Acanthocystis, Heterophrys, Raphidocystis |
| Copromyxids | slime mould | 2 genera |
| Desmothoracids | heliozoon-like | e.g. Clathrulina |
| Entamoebidae | amoebae, mitochondria absent | A few genera only, e.g. Entamoeba |
| Excavates | mostly flagellates, but amoeboid and slime mould organisms occur within the Heterolobosea | e.g. Naegleria, Acrasis (acrasid slime moulds) |
| Fonticula | slime-mould | Isolated genus |
| Granuloreticulosa | amoebae with thin, stiff and anastomosing pseudopodia, most (Foraminifera) with a single or many chambered shell. | Very diverse, with a substantial fossil record, e.g. Reticulomyxa, Lagena, Globigerina, Elphidium. |
| Gymnophrea | several genera of amoebae, thin pseudopodia, with short inactive flagellar stumps | Gymnophrys, Barkowia |
| Gymnosphaerida | a type of heliozoon | Gymnosphaera, Hedraiophrys, Actinocoryne |
| Komokiacea | very large marine amoebae | About 12 genera |
| Nucleariidae | with thin unbranching pseudopodia | Nuclearia, Vampyrellidium, Pinaciophora, Pompholyxophrys, Rabdiophrys |
| Pelobionts | several genera of amitochondriate flagellates, but one genus is often considered as an amoeba | Pelomyxa |
| Phaeodarea | a type of radiolaria, common | About 100 genera |
| Polycystinea | a type of radiolaria, common | Diverse |
| Ramicristates | this group contains most of the non-actinopod amoebae | Naked amoebae, testate amoebae, dictyostelid and myxomycete slime moulds, diverse, e.g. Hyperamoeba, Gromia, Acanthamoeba |
| Sticholonche | genus of heliozoa with mobile arms | Single genus, two species |
| Vampyrellids | fungus- and alga-eating amoebae | e.g. Vampyrella |
| Xenophyophores | large marine amoebae | About 15 genera, 50 species |
Genera of amoebae for which no clear identity has been developed (after Patterson, 1999):
|
|
|
|
|