



Bombinatoridae
Bombinas and barbourulas
David Cannatella




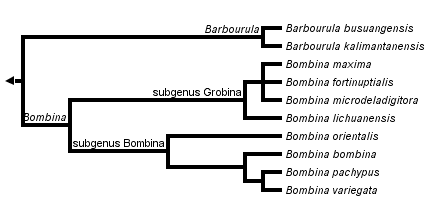


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
This family includes only two genera, Barbourula and Bombina. Barbourula occurs in the Philippine Islands (Barbourula busuangensis) and Borneo (Barbourula kalimantanensis). The approximately eight species of Bombina are found in Europe, western Asia, China, Vietnam, and Korea.
Bombina are warty, aquatic toads, and tend to be gregarious. The vocal behavior of some Bombina is unusual in that the call is produced during inhalation rather than exhalation as in other frogs. They lay pigmented eggs in ponds. The tadpoles have beaks, two upper and three lower rows of denticles, and a median spiracle (typical Orton Type 3).
Species of Bombina have bright red or yellow mottling on the venter. The German word Unke means toad. Bombina exhibit an "unken reflex" when bothered. The animal will arch its back and limbs to expose the bright belly, and may turn over on its back. This acts as a warning to predators. Not surprisingly, the skin toxins of Bombina are distasteful.
Barbourula are not as brightly colored but have webbed fingers in addition to webbed toes. Tadpoles of Barbourula are unknown. Both genera have flattened bodies.
Fossil Bombina are known from the Pliocene to the Pleistocene; there are no fossils of Barbourula.
Discussion of Phylogenetic Relationships
Morphological synapomorphies of Bombinatoridae include an expanded flange of the quadratojugal and the presence of endochondral ossifications in the hyoid plate; these are the same as parahyoid bones, which are also present (Cannatella, 1985; Clarke, 1987).
Yu et al. (2007) estimated the phylogeny of most of the species of Bombina using several mitochondial genes; they recovered a largely European clade (subgenus Bombina) and a largely Asian clade (subgenus Grobina).
Relationships to Outgroups
Prior to 1985, Bombina and Barbourula were generally placed in the Discoglossidae, with Alytes and Discoglossus, although the dissimilarity of Alytes and Discoglossus, on one hand, and Bombina on the other has often been noted (e.g., Lanza et al., 1976).
Cannatella (1985) presented a phylogenetic analysis indicating Discoglossidae was paraphyletic. Ford and Cannatella (1993) formalized this by defining Bombinatoridae to be the node-based name for the most recent common ancestor of Bombina and Barbourula, and all its descendants.
In contrast to Cannatella (1985), Hay et al. (1995) found Bombina orientalis and Discoglossus pictus to be closest relatives; this supports the monophyly of Discoglossidae in the traditional sense. Roelants and Bossuyt (2005) found a similar result, and continued recognition of Bombinatoridae and Discoglossidae.
Griffiths (1963) stated that the diagnostic morphological feature of Discoglossidae sensu lato (including Bombinatoridae and Discoglossidae) is a triradiate sternum. This type of sternum is also present in Leiopelma.
References
Cannatella, D. C. 1985. A phylogeny of primitive frogs (archaeobatrachians). Ph.D. Dissertation, The University of Kansas, Lawrence.
Clarke, B. T. 1987. A description of the skeletal morphology of Barbourula (Anura: Discoglossidae), with comments on its relationships. J. Nat. Hist. 21:879-891.
Ford, L. S., and D. C. Cannatella. 1993. The major clades of frogs. Herp. Monogr. 7:94-117.
Griffiths, I. 1963. The phylogeny of the Salientia. Biological Reviews 38:241-292.
Hay, J. M., I. Ruvinsky, S. B. Hedges, and L. R. Maxson. 1995. Phylogenetic relationships of amphibian families inferred from DNA sequences of mitochondrial 12S and 16S ribosomal RNA genes. Mol. Biol. Evol. 12(5):928-937.
Lanza, B., J. M. Cei, and E. G. Crespo. 1976. Further immunological evidence for the validity of the family Bombinidae (Amphibia, Salientia). Monit. Zool. Ital. (NS) 10:311-314.
Roelants, K., and F. Bossuyt. 2005. Archaeobatrachian paraphyly and Pangaean diversification of crown-group frogs. Syst. Biol. 54(1):111-126.
Yu, G., J. Yang, M. Zhang, and D. Rao. 2007. Phylogenetic and systematic study of the genus Bombina (Amphibia: Anura: Bombinatoridae): new insights from molecular data. Journal of Herpetology 41(3):365-377.
Title Illustrations

| Scientific Name | Bombina orientalis |
|---|---|
| Image Use |
 This media file is licensed under the Creative Commons Attribution-ShareAlike License - Version 3.0. This media file is licensed under the Creative Commons Attribution-ShareAlike License - Version 3.0.
|
| Copyright |
© 1995 David Cannatella

|

| Scientific Name | Bombina orientalis |
|---|---|
| View | ventral |
| Image Use |
This media file is licensed under the Creative Commons Attribution-ShareAlike License - Version 3.0.
|
| Copyright |
© 1995 David Cannatella
|

| Scientific Name | Bombina bombina |
|---|---|
| Location | Romania |
| Specimen Condition | Live Specimen |
| Identified By | Horia Bogdan |
| Copyright |
© Horia Bogdan
|

| Scientific Name | Bombina bombina |
|---|---|
| Location | Romania |
| Specimen Condition | Live Specimen |
| Identified By | Horia Bogdan |
| Copyright |
© Horia Bogdan
|
About This Page
If you are interested in authoring or co-authoring the page for this taxon, or some part of it (even a species), contact David Cannatella.
David Cannatella
University of Texas, Austin, Texas, USA
Correspondence regarding this page should be directed to David Cannatella at
Page copyright © 1995 David Cannatella
 Page: Tree of Life
Bombinatoridae. Bombinas and barbourulas.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Bombinatoridae. Bombinas and barbourulas.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 29 November 2008
Citing this page:
Cannatella, David. 2008. Bombinatoridae. Bombinas and barbourulas. Version 29 November 2008 (under construction). http://tolweb.org/Bombinatoridae/16971/2008.11.29 in The Tree of Life Web Project, http://tolweb.org/

