




Hydrophiloidea
Michael S. Caterino




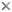
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Hydrophiloidea is a moderately large superfamily, containing about 6500 species distributed throughout the world (Hansen, 1991; Newton, 1991; Mazur, 1997). They are very diverse in habits. Approximately one-fourth of the superfamily is strictly aquatic, as larvae and adults, while the remainder is terrestrial. The larvae of virtually all known species are predaceous, and approximately two-thirds of the species are also predaceous as adults. But adults of most Hydrophilidae and Sphaeritidae are saprophagous.
Characteristics
Adult Hydrophiloidea are very diverse in body form. The one essentially constant character is the terminal 3 (rarely 5) segments of the antennae forming a distinct, pubescent club. The segment preceding this club is also usually cup-like. The group is more easily characterized by their larvae. Most Hydrophiloid larvae have desclerotized abdomens and articulated urogomphi; they are prognathous, with generally falcate mandibles (lacking molar region), and they have the labrum (and clypeus) indistinguishably fused to the head capsule forming a toothed "nasale."
Discussion of Phylogenetic Relationships
The monophyly of Hydrophiloidea is generally accepted (Lawrence & Newton, 1982, 1995), although not entirely uncontroversial. Characters supporting hydrophiloid monophyly are most apparent in the larval stages. Of the characters listed above, prognathy and the modifications of the clypeolabral region (fusion of these scelerites to the frons) are apparent synapomorphies. Additional possible synapomorphies include the form of the antennae and maxillary palpus. However, adult synapmorphies are somewhat more obscure. Hansen (1997) cites three possible characters: atrophied spiracles of abdominal segment 7, male tergite 9 with a sclerotized dorsal bridge, and head with fimbriate epistomal lobe. An old, largely dead, controversy placed Hydraenidae as the sister group to Hydrophilidae, rather than in the Staphylinoidea. The two groups both have aquatic adults, and share numerous characteristics related to this habit. But larvae of Hydraenidae have little in common with those of Hydrophilidae, and this question is considered settled. Recently, some molecular data have suggested some unconventional relationships, placing the Hydrophilidae within Scarabaeoidea (Hawks, unpublished). However, these data are still being analyzed (Hawks, pers. comm.), so it is impossible to evaluate this suggestion.
There is no question about the monophyly of each the four families recognized here. Hydrophilidae is frequently divided into as many as 6 families (e.g. Hansen, 1991), but these are treated as subfamilies here. Relationships among the families of Hydrophiloidea have not received tremendous attention, but the relationships presented here have been long suspected, and have recently found support by morphology (Hansen, 1997) and molecules (Caterino & Vogler, in press).
The monophyly of Sphaeritidae + Synteliidae + Histeridae is supported by several characters, including truncate elytra, compact antennal club of the adults, complete invagination of abdominal segment 8 into the abdomen, scooplike coxites of the ovipositor, lack of stemmata in the larvae, and numerous other features (see also Hansen, 1997; Lawrence and Newton, 1982; Newton, 1991; Slipinski & Mazur, 1999; Caterino & Vogler, in press). Ecology is also entirely consistent with these relationships. Like Hydrophilidae, Sphaeritidae is known to be saprophagous in the adult stage (Nikitsky, 1976; Löbl, 1996; Newton, 2001), while Synteliidae and Histeridae have become primarily predaceous in all life stages (Mamayev, 1974; Newton, 1991). The sister group relationship between Synteliidae and Histeridae is also supported by several morphological characters including concealment of the protrochanters (adults), sclerotization of the abdominal tergites from at least tergite 4 and back, and loss of the cardo of the larval maxilla (Ohara, 1995; Hansen, 1997; Slipinski and Mazur, 1999).
References
Archangelsky, M. 1998. Phylogeny of Hydrophiloidea using characters from adult and preimaginal stages. Systematic Entomology 23: 9-24.
Caterino, M. S. and A. P. Vogler. in press. The phylogeny of the Histeroidea. Cladistics.
Beutel, R. G. 1994. Phylogenetic analysis of Hydrophiloidea based on characters of the head of adults and larvae. Koleopterologische Rundschau 64: 103-131.
Beutel, R. G. 1999. Morphology and evolution of the larval head of Hydrophiloidea and Histeroidea. Tijdschrift voor Entomologie 142: 9-30.
Hansen, M. 1991. The hydrophiloid beetles. Phylogeny, classification and a revision of the genera. Biologiske Skrifter 40: 1-368.
Hansen, M. 1995. Evolution and classification of the Hydrophiloidea - A systematic review. Pages 321-353 in Biology, Phylogeny, and Classification of the Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson (J. Pakaluk, and S. A. Slipinski, eds.). Muzeum i Instytut Zoologii PAN, Warsaw.Hansen, M. 1997. Phylogeny and classification of the staphyliniform beetle families. Biologiske Skrifter 48: 1-339.
Lawrence, J. F., and A. F. Newton. 1982. Evolution and classification of beetles. Annual Review of Ecology and Systematics 13:261-290.
Lawrence, J. F. and A. F. Newton. 1995. Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). In J. Pakaluk and S. A. Slipinsky (eds), Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). Muzeum i Instytut Zoologii PAN, Warsaw, pp. 779-1006.
Mamayev, B. M. 1974. The immature stages of the beetle Syntelia histeroides Lewis in comparison with certain Histeridae. Entomological Review 53: 98-101.
Newton, A. F. 1991. Agyrtidae-Silphidae (pp. 324-341); Pselaphidae (pp. 353-355); Sphaeritidae-Histeridae (pp. 341-352). In F. W. Stehr (ed) Agyrtidae-Silphidae (pp. 324-341); Pselaphidae (pp. 353-355); Sphaeritidae-Histeridae (pp. 341-352). Kendall/Hunt, Dubuque.
Newton, A. F. 2001. Sphaeritidae. In R. H. Arnett and M. C. Thomas (eds), American Beetles. CRC Press, Boca Raton.
Nikitsky, N. B. 1976b. On the morphology of a larva Sphaerites glabratus and the phylogeny of Histeroidea. Zoologicheskii Zhurnal 55: 531-537.
Ohara, M. 1994. A revision of the superfamily Histeroidea of Japan. Insecta Mastumurana (N.S.) 51: 1-283.
Slipinski, S. A., and S. Mazur. 1999. Epuraeosoma, a new genus of Histerinae and phylogeny of the family Histeridae. Annales Zoologici (Warszawa) 49:209-230.
Title Illustrations

| Scientific Name | Anapleus cyclonotus |
|---|---|
| Location | India |
| Specimen Condition | Dead Specimen |
| Copyright | © 2002 |

| Scientific Name | Saprinus discoidalis |
|---|---|
| Location | California |
| Specimen Condition | Dead Specimen |
| Copyright | © 2002 |

| Scientific Name | Tropisternus |
|---|---|
| Specimen Condition | Dead Specimen |
| Copyright | © 2002 |
About This Page
Santa Barbara Museum of Natural History
2559 Puesta del Sol Rd.
Santa Barbara, CA 93105 USA
Correspondence regarding this page should be directed to Michael S. Caterino at
Page copyright © 2002
- First online 07 March 2002
Citing this page:
Caterino, Michael S. 2002. Hydrophiloidea. Version 07 March 2002 (under construction). http://tolweb.org/Hydrophiloidea/9075/2002.03.07 in The Tree of Life Web Project, http://tolweb.org/
