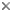Pterotracheoidea
Heteropoda , heteropods, sea elephants
Roger R. Seapy



- Atlantidae Rang, 1829
- Pterotracheidae Rafinesque, 1814
- Carinariidae Blainville 1818
Introduction
The heteropods are a group of pelagic snails (Class Gastropoda) that are found in moderate to low abundances, primarily in tropical to subtropical latitudes. Among the gastropods, they have three striking adaptations to the open ocean environment. The bodies and shells are largely transparent; only the buccal mass, eyes and viscera are opaque. The transparent body allows one to view internal structures in the head region, such as the eyes, statocysts and central nervous system.
Figure. Transparent head region of Pterotrachea hippocampus. ©
The foot, which in bottom-dwelling snails is the sole-like structure used for crawling along the bottom, primarily forms a laterally-compressed swimming fin. The animal swims "upside-down" with the fin directed upward. The eyes are large and image-forming. They are active predators that visually locate their prey and capture them by protruding their radula (having elongate, sickle-shaped teeth) from the mouth at the tip of the proboscis. The presence øf a mobile, trunk-like proboscis is responsible for their common name, "sea elephants."
Diagnosis
Gastropod molluscs with:- Paired image-forming eyes with large, spherical lenses.
- Laterally-flattened ventral swimming fin.
Characteristics
- Eyes
- Eye forward-looking, with large, spherical lens.
- Eye with basal, ribbon-like retina.
- Eye shape, viewed from dorsal aspect, generally tubular to triangular.
- Wall of eye partially to completely pigmented; in the former case, the unpigmented region forms a "window".
- Eye enclosed in gelatinous capsule. Click on an image to view larger version & data in a new window

Figure. Dorsal view, left eye of Pterotrachea hippocampus, small animal (about 20 mm body length), Hawaiian waters. Eye with narrowly triangular shape. © 2005 R. Seapy.
- Foot
- Foot laterally compressed into swimming fin.
- Swimming fin usually with a ventral to posteroventral sucker. Click on an image to view larger version & data in a new window

Figure. Swimming fin with posteroventral sucker in male Carinaria japonica, viewed from the right side. copyright © 2005 R. Seapy.
- In atlantids the sucker is much larger than in the other two families (it serves the function of holding prey for ingestion) and the posterior margin of the fin forms an opercular lobe, to which the operculum is attached (see Atlantidae page and title illustration).
- Proboscis and radula
- Proboscis muscular and mobile, normally extending anteriorly from the trunk.
- Proboscis bears terminal buccal mass with radula. Click on an image to view larger version & data in a new window


Figure. Dorsal view of head and proboscis (left) and buccal mass with radula (right) of female Pterotrachea scutata, Hawaiian waters. Copyright © 2005 R. Seapy.
- Radula taenioglossate with a tooth formula of 2-1-R-1-2; i.e., each tooth row consists of two marginal teeth and one lateral tooth on either side of the central rachidian tooth. Rachidian tooth morphology differs between families. Lateral and marginal teeth with elongated shafts and hook-shaped ends. Click on an image to view larger version & data in a new window


Figure. Dorsal views of an atlantid radula. Left - Photomicrograph using dissection microscope. Right - Photomicrograph using scanning electron microscope. Photographs copyright © 2005 R. Seapy (left) and G. Richter (right).
- Radular tooth shapes change during ontogenesis, with the result that position on the radula of the tooth row can change the appearance of the teeth (Richter, 1961, 1963). Morphogenetic changes in the teeth continue for a large part of radular growth in the Atlantidae, while in the other two families change is restricted to the late larval and early post-larval stages.
- Two markedly different types of radulae are distinguished in the heteropods (reviewed in Richter and Seapy, 1999). In Type I radulae, the number of tooth rows increases continuously with growth, with the result that all stages of change in tooth shape are represented in mature animals and that teeth produced first on the radula are not cast off the anterior end of the radula after they are no longer of use in prey capture. In Type II radulae, teeth are cast off the anterior end of the radula once the radula has reached its specific number of tooth rows. Thus, the radula has a limited number of tooth rows. Type II radulae are seen in all Carinariidae and Pterotracheidae, while in the Atlantidae both types are seen (13 species have Type I and 8 species have Type II radulae).
- Shell
- Coiled shell present in larvae of all three families.
- Larval shell retained in adult Atlantidae and Carinariidae, but is cast off at metamorphosis in Pterotracheidae.
- When present, adult shell with keel, developed to varied degrees.
- Adult shell calcareous (aragonitic) in adult Carinariidae, but in Atlantidae shell and keel can be calcareous (Atlanta), composed exclusively of conchiolin (a cartilaginous-like material; Oxygyrus), or a combination of the two (shell calcareous and keel conchiolin; Protatlanta).Click on an image to view larger version & data in a new window


Figure. Left - Right side of transparent shell in Carinaria galea. Well-developed keel extends to the right from the anterior edge of the shell. Shell surrounds visceral mass and gills, all of which is attached to the posterior part of the trunk by a stalk. Right - Right side of Atlanta peroni with transparent shell, Hawaiian waters. Prominent keel surrounds the outermost shell whorl. © 2005 R. Seapy.
Comments
The Pterotracheidae includes three families that can be most readily distinguished by the following characters:
| Family | Adult shell | Body can retract into shell | Adult body size | Location of swimming fin |
|---|---|---|---|---|
| Atlantidae | present | yes | microscopic (shell diameter < 1 cm) | opposite visceral mass |
| Carinariidae | present | no | macroscopic (adult body length > ca. 2 cm) | opposite visceral mass |
| Pterotracheidae | absent | not applicable | macroscopic (adult body length > ca. 2 cm) | between visceral mass and head |
Life History
Heteropods are dioecious (separate sexes) and exhibit sexual dimorphism. Males have a prominent penis and penial appendage (function unknown) and the fin sucker in pterotracheids is present only in males. Also, there are sexual differences in radular morphology in some atlantids.
Male heteropods package sperm into spermatophores. Sperm are transferred to the penis by an external ciliated groove where they are packed into spermatophores. Spermatophores are transferred to females during copulation (mating behavior has only been observed in Pterotrachea hippocampus; Harbison in Lalli and Gilmer, 1989). Fertilized eggs are usually layed in mucoid egg strings which eventually break free from the female, except in Firoloida desmaresti which has a permanently attached tubular filament which holds the developing embryos (Owre, 1964).


Figure. Left - Side view of posterior region of trunk of Carinaria japonica, male, Hawaiian waters. Penis and penial appendage lie parallel to each other; penis contains brown spermatophore. Right - Side view of visceral nucleus in Pterotrachea coronata, female, Hawaiian waters. The egg string is released from the oviduct opening, which is surrounded by transparent "lips". Photographs copyright © 2005 R. Seapy.
All heteropods have a free-swimming, planktotrophic veliger larva. Larvae hatch from the egg within a few days of fertilization. The veliger possesses a dextrally (right-hand) coiled, calcareous shell and a flexible, chitinous operculum. The larval velum (ciliated swimming and feeding organ) is initially small and bilobed. With growth each lobe forms into two (pterotracheids) or three (atlantids and carinariids) long and slender lobes. Following metamorphosis the larval shell is retained as the protoconch of the adult shell (in atlantids and carinariids) or is cast off (in pterotracheids, which lack an adult shell). The larval operculum is either retained (atlantids) or cast off (carinariids and pterotracheids). The duration of the larval stage is not known, nor is age or life span of the adults. The largest heteropods are the carinariids (to 500 mm in Carinaria cristata) followed by the pterotracheids (to 330 mm in Pterotrachea coronata) (Lalli and Gilmer, 1989).

Figure. Left - Oral view of veliger larva of Carinaria lamarcki with six slender, ciliated velar lobes. Drawing from Thiriot-Quievrèux (1973). Right - Late veliger larva of Oxygyrus keraudreni, shell diameter about 1.0 mm, from Hawaiian waters. Velar lobes (light violet color) retracted into shell; head, proboscis and opercular lobe extend from shell. Photograph copyright © 2005 R. Seapy.
Natural History
All heteropods swim with their ventral side directed upward and the dorsal side, with the shell (when present) and visceral mass downward. Steady swimming is by undulations of the fin. In atlantids, a considerable side-to-side body motion is induced by the large fin, although the shell and keel act to partially offset the fin's movements. The large, elongate bodies of the carinariids and pterotracheids greatly dampen the side-to-side body motion. In the latter two groups, and especially in the pterotracheids, flexion of the trunk and tail are used in accelerated swimming during pursuit of prey or evasion of predators. The bodies of pterotracheids are the most elongate and streamlined of the three families, and they are the fastest swimmers.
The atlantids are negatively buoyant. During the day they must swim to maintain position. At night, however, they secrete strands of buoyant mucus, up to 0.5 m long, from which they are suspended (Lalli and Gilmer, 1989; Newman, 1990). Carinariids and pterotracheids have been observed to float motionless without sinking (Lalli and Gilmer, 1989). Their neutral buoyancy results from the large amounts of gelatinous tissue, mostly in the trunk and tail, that is made positively-buoyant by ion regulation; i.e., by replacing heavier sulfate ions by lighter chloride ions (up to 75% in Pterotrachea coronata) (Denton and Shaw, 1961).
Heteropods are carnivores that feed during the day, locating their prey visually. The eyes are unusual because they have a narrow, strip-like retina. Image formation is accomplished by scanning-eye movements. In atlantids, the only family studied to date, the retina is rotated through a 90° arc, from downward vertical to horizontal (Land, 1982). Presumably, prey are located beneath the animal by light reflected off the prey's body. In all heteropods, prey are captured by elongate, hooked radular teeth (described above). In the atlantids, the fin sucker is large and is used to hold the prey while pieces of tissue are torn off and ingested. In the carinariids and pterotracheids prey are ingested whole. The fin sucker is much smaller in these two groups and does not appear to be used in feeding. It is directed ventrally to posteroventrally and is spatially separated from the mouth; its primary or sole function is presumably to hold males and females together during mating.
Heteropods feed on a variety of zooplanktonic prey. Atlantids feed preferentially on other gastropods, especially shelled pteropods (Richter, 1968; Newman, 1990), and carinariids feed selectively on soft bodied prey (e.g., salps, doliolids and chaetognaths in Carinaria japonica; Seapy, 1980). Little is known of feeding in pterotracheids.
Defense against predators is based largely on transparency, although the buccal mass, pigmented region of the eyes, and the visceral mass (or visceral nucleus) are opaque. Pterotrachea uses reflectivity and behavioral orientation to reduce the visibility of these structures to upward-searching predators (Seapy and Young, 1986).
References
Denton, E. J. and T. I. P. Shaw. 1961. The buoyancy of gelatinous marine animals. Journal of Physiology, London 161: 14P-15P (Proceedings).
Lalli, C. M. and R. W. Gilmer. 1989. Pelagic snails. The biology of holoplanktonic gastropod snails. Stanford: Stanford University Press. 259 pp.
Land, M. F. 1982. Scanning eye movements in a heteropod mollusc. Journal of Experimental Biology 96: 427-430.
Newman, L. 1990. The taxonomy, distribution and biology of Atlanta gaudichaudi Souleyet, 1852 (Gastropoda, Heteropoda) from the Great Barrier Reef. American Malacolological Union Bulletin 8: 85-94.
Newman, L. 1998. Superfamily Carinarioidea, pp. 804-808. In: P. L. Beesley, G. L. B. Ross and A. Wells (eds), Mollusca: the southern synthesis. The Fauna of Australia. Vol. 5, Pt. B. Melbourne: CSIRO Publ.
Owre, H. B. 1964. Observations on development of the heteropod molluscs Pterotrachea hippocampus and Firoloida desmaresti. Bulletin of Marine Science 14: 529-538.
Richter, G. 1961. Die Radula der Atlantiden (Heteropoda, Prosobranchia) und ihre Bedeutung fur die Systematik und Evolution der Familie. Zeitschrift f?r Morphologie und ?kologie der Tiere 50: 163-238.
Richter, G. 1963. Untersuchungen zur Morphogenese der Gastropodenradula. Veroffentlichungen des Instituts fur Meeresforschung in Mremerhaven, Drittes meeresbiologisches Symposion 3: 142-152.
Richter, G. 1968. Heteropoden und Heteropodenlarven im Oberfl?chenplankton des Golfs von Neapel. Pubblicazioni della Stazione Zoologica di Napoli 36: 346-400.
Richter, G. 1974. Die Heteropoden der "Meteor"-Expedition in den Indischen Ozean, 1964/65. "Meteor" Forschungs-Ergebnisse (D), 17: 55-78.
Richter, G. and R. R. Seapy. 1999. Heteropoda, pp. 621-647. In: D. Boltovskoy (ed.), South Atlantic Zooplankton. Leiden: Backhuys Publ.
Seapy, R. R. 1980. Predation by the epipelagic heteropod mollusk Carinaria cristata forma japonica. Marine Biology 60: 137-146.
Seapy, R. R. and R. E. Young. 1986. Concealment in epipelagic pterotracheid heteropods (Gastropoda) and cranchiid squids (Cephalopoda). Journal of the Zoological Society of London (A) 210: 137-147.
Seapy, R. R., C. M. Lalli and F. E. Wells. 2003. Heteropoda from western Australian waters, pp. 513-546. In: F. E. Wells, D. I. Walker and D. S. Jones (eds.), The Marine Flora and Fauna of Dampier, Western Australia. Perth: Western Australia Museum.
Spoel, S. van der. 1976. Pseudothecosomata, Gymnosomata and Heteropoda (Gastropoda). Utrecht: Bohn, Scheltema & Holkema. 484 pp.
Spoel, S. van der, L. Newman and K. W. Estep. 1997. Pelagic molluscs of the world. World Biodiversity Data Base CD-ROM Series. Amsterdam: Expert Center for Taxonomic Identification (ETI).
Tesch, J. J. 1949. Heteropoda. Dana Report, No. 34. 53 pp.
Thiriot-Qui?vreux, C. 1973. Heteropoda. Oceanography and Marine Biology, Annual Review 11: 237-261.
Title Illustrations

| Scientific Name | Oxygyrus keraudreni |
|---|---|
| Location | Hawaiian waters |
| Specimen Condition | Live Specimen |
| Sex | Female |
| Life Cycle Stage | adult |
| View | right side |
| ToL Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2005

|

| Scientific Name | Carinaria japonica |
|---|---|
| Location | Hawaiian waters |
| Specimen Condition | Live Specimen |
| Sex | Male |
| Life Cycle Stage | adult |
| View | right side |
| ToL Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2005
|

| Scientific Name | Pterotrachea coronata |
|---|---|
| Location | Hawaiian waters |
| Specimen Condition | Live Specimen |
| Sex | Female |
| Life Cycle Stage | adult |
| View | right side |
| ToL Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2005
|
About This Page
California State University, Fullerton, California, USA
Correspondence regarding this page should be directed to Roger R. Seapy at
Page copyright © 2005
 Page: Tree of Life
Pterotracheoidea . Heteropoda , heteropods, sea elephants.
Authored by
Roger R. Seapy.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Pterotracheoidea . Heteropoda , heteropods, sea elephants.
Authored by
Roger R. Seapy.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 15 February 2005
- Content changed 21 September 2008
Citing this page:
Seapy, Roger R. . 2008. Pterotracheoidea . Heteropoda , heteropods, sea elephants. Version 21 September 2008 (under construction). http://tolweb.org/Pterotracheoidea/27801/2008.09.21 in The Tree of Life Web Project, http://tolweb.org/