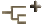The phylogeny of early groups of amniotes has been in a state of flux in the last few years, but the first phylogeny based on a data matrix was published in the eighties (Gauthier et al., 1988). The origins of mammals and saurians from early synapsids and early diapsids, respectively, has been relatively non-controversial, but the origin of turtles has been problematic. The most widespread view, at least until the last few years, was that the closest extinct relatives of turtles were the captorhinids (Gaffney & Mc Kenna, 1979; Gaffney & Meylan, 1988; Gaffney, Meylan, & Wyss, 1991; Gauthier et al., 1988). The study of Gauthier et al. (1988) is probably the best documented study supporting close affinities between captorhinids and turtles:
======================================= Synapsida
|
| ===== Millerettidae
| ===|
| ===| ===== Pareiasauria
| ==parareptiles=| |
| | | ======== Procolophonidae
| | |
=====| | =========== Mesosauridae
==Sauropsida=|
| ==== Captorhinidae
| ==Anapsida=|
| | ==== Testudines
==Reptilia=|
| ||| Protorothyrididae
==Romeriida=|
=== Diapsida
This study is also significant in giving the first phylogenetic definitions for the names of many amniote taxa. In this tree, parareptiles are not closely related to turtles, but Gauthier et al. (1988) were not confident in this part of their tree (which is why they did not erect a formal Parareptilia).
Laurin & Reisz (1995) suggested that many of the taxa that Gauthier et al. (1988) considered parareptiles represented early relatives of turtles. Furthermore, Laurin & Reisz (1995) suggested that mesosaurs were the closest known relatives of reptiles:
==================================================== Synapsida
|
| ======================================= Mesosauridae
| |
| | ============= Millerettidae
| | |
| | | ========== Pareiasauria
| | ==Parareptilia=| |
=====| | | ===| ======= Procolophonidae
==Sauropsida=| | ===|
==Reptilia=| ======= Testudines
|
| =============== Captorhinidae
| |
==Eureptilia=| ||| Protorothyrididae
==Romeriida=|
=== Diapsida
This study used a terminology derived from Olson (1947). Unfortunately, this terminology includes some junior synonyms (according to the rules of priority suggested by de Queiroz & Gauthier in 1990, 1992, and 1994) of the taxa defined by Gauthier et al. (1988) and the terminology used in the main phylogeny (in the page on Amniota) corrects this.
Lee (1995, 1996) studied in detail the phylogeny of the presumed relatives of turtles, and argued that pareiasaurs are the closest known relatives of turtles, rather than procolophonids (as argued by Laurin & Reisz, 1995). Furthermore, he believed that pareiasaurs are the stem-group of turtles (some pareiasaurs are more closely related to turtles than to other pareiasaurs). His phylogeny is as follows:
============================================================= Nyctiphruretia
|
| ===================== Owenettidae
| ==Procolophonoidea=|
| | ===================== Procolophonids
| |
=====| | ======================== Lanthanosuchids
==Procolophoniformes=| |
| | ====== Sclerosaurus
==Hallucicrania=| |
==Pareiasauroidea=| ||| Pareiasauria
===|
=== Testudines
All the studies mentioned above agree that turtles are the sister-clade of saurians among extant amniotes. However, the old idea that turtles are closely related to placodonts (Broom, 1924) has been revived recently (Rieppel, 1994, 1995; Rieppel & deBraga, 1996; deBraga & Rieppel, 1997). If this theory is accurate, turtles are saurians and are more closely related to lepidosaurs than to archosaurs.
================== Captorhinidae
|
| ============ Pareiasauria
| ===|
| | ============ Procolophonidae
| |
| | ============ Araeoscelidia (a Permo-Carboniferous group of diapsids)
=====| | |
===| | ====== Lepidosauria
| | |
===| ===| === Sauropterygia (a Mesozoic group of aquatic diapsids)
===| ===|
| === Testudines
|
========= Archosauromorpha
Amniote classification and characters supporting amniote phylogeny
An exhaustive list of autapomorphies of various amniote taxa cannot be provided here, but a sample of these characters can be given. The list given below assumes that the phylogeny presented at the beginning of the page on Amniota is essentially correct. Phylogenetic definitions, when available, are also given for the main taxa. Amniote classification can be summarized as such:
Amniota
Synapsida
Sauropsida
Mesosauridae
Reptilia
Anapsida
Millerettidae
Unnamed
Unnamed
Acleistorhinus
Lanthanosuchidae
Unnamed
Macroleter
Nyctiphruretia
Procolophonia
Pareiasauria
Procolophonoidea
Testudines
Romeriida
Captorhinidae
Unnamed
"Protorothyrididae"
Diapsida
Parts of the phylogeny are poorly documented (most of the unnamed clades) and will not be discussed below. Furthermore, if the suggestion of Modesto (1999) that mesosaurs are anapsids is accepted, this classification and the list of apomorphies will have to be modified.
Amniota is defined as "the most recent common ancestor of extant mammals and reptiles, and all its descendants" (Gauthier et al., 1988). It is divided into two stem-based taxa: Synapsida (mammals and their extinct relatives) and Sauropsida (reptiles and their fossil relatives). The autapomorphies of Amniota are listed in the "Characteristics" section of the Tree of Life page.
Sauropsida is defined as "reptiles plus all other amniotes more closely related to them than they are to mammals" (Gauthier, 1994). The characters supporting Sauropsida include the following:
- Presence of a single coronoid. Synapsids and the diadectomorph Limnoscelis have two coronoid elements. The coronoids are bones on the dorsomedial surface of the lower jaw.
- Supinator process parallel to humeral shaft and separated from it by a groove. In synapsids and diadectomorphs, the supinator process is strongly angled relative to the shaft.
- Presence of a single pedal centrale. Two centralia were present in the tarsus (ankle) of synapsids and diadectomorphs.
Reptilia is defined as "the most recent common ancestor of extant turtles and saurians, and all of its descendants" (Gauthier et al., 1988). Characters supporting Reptilia include:
- Tabular small. The tabular (a bone on the posterolateral corner of the skull table) of early synapsids and diadectomorphs is large, but reptiles have only a small tabular, when it is present.
- Suborbital foramen present (Fig. 1B-D). The suborbital foramen is a small hole near the lateral edge of the palate, between the pterygoid, palatine, and ectopterygoid (or jugal, when the ectopterygoid is absent). This structure was not found in early synapsids and diadectomorphs (Fig. 1A).
- Supraoccipital anterior crista present. The supraoccipital of mesosaurs, synapsids, and diadectomoprhs lacks anterior parasagittal flanges. The supraoccipital of reptiles has a paired anterior parasagittal flange called an anterior crista.
- Supraoccipital plate narrow. The supraoccipital is a bone in the back of the braincase. The supraoccipital plate of mesosaurs, synapsids, and diadectomorphs is broad and extends farther laterally than the postparietal.

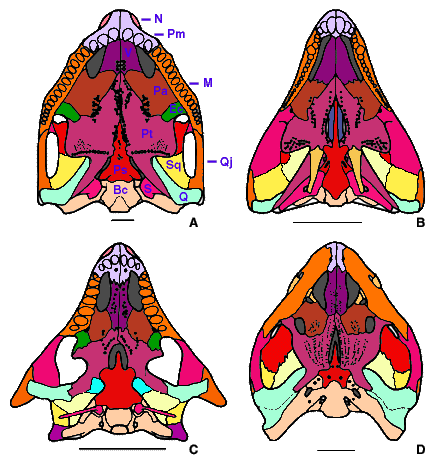
Figure 1. Amniote skulls in palatal view. A, Cotylorhynchus, a Lower Permian synapsid; B, Captorhinus, a Lower Permian romeriid; C, Procolophon, a Triassic anapsid; D, Proganochelys, the oldest known turtle (Upper Triassic). Redrawn from A, Laurin & Reisz (1995); B, Heaton (1979); C, Carroll & Lindsay (1985); and D, Gaffney (1990). Abbreviations: Bc, braincase; Ec, ectopterygoid; M, maxilla; N, nasal; Pa, palatine; Pm, premaxilla; Ps, parasphenoid; Pt, pterygoid; Q, quadrate; Qj, quadratojugal; S, stapes; Sq, squamosal; V, vomer.
Anapsida is defined as "extant turtles, and all other extinct taxa that are more closely related to them than they are to other [extant] reptiles" (Gauthier et al., 1988). Characters supporting Anapsida include:
- Foramen orbito-nasale enclosed between prefrontal, lacrimal, and palatine. There is no foramen between these bones in diapsids, synapsids, and diadectomorphs.
- Anterior lateral maxillary foramen distinctly larger than other foramina. In romeriids, mesosaurs, synapsids, and diadectomorphs, several small foramina (holes) may be present on the lateral surface of the maxilla, but none of these foramina is much larger than the others.
- Quadratojugal expanded dorsally (Fig. 2C, D). The quadratojugal of romeriids, mesosaurs, synapsids, and diadectomorphs is a low bone narrowly exposed on the lateral surface of the cheek (Fig. 2A, B).
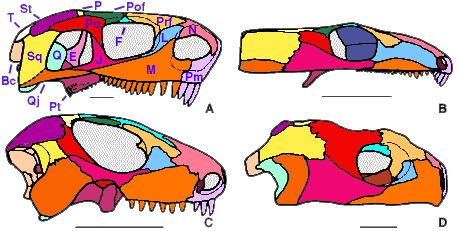
Figure 2. Amniote skulls in lateral view. A, Cotylorhynchus, a Lower Permian synapsid; B, Captorhinus, a Lower Permian romeriid; C, Procolophon, a Triassic anapsid; D, Proganochelys, the oldest known turtle (Upper Triassic). Redrawn from A, Laurin & Reisz (1995); B, Heaton (1979); C, Carroll & Lindsay (1985); and D, Gaffney (1990). Abbreviations: Bc, braincase; E, epipterygoid; F, frontal; L, lacrimal; M, maxilla; N, nasal; P, parietal; Pm, premaxilla; Po, postorbital; Pof, postfrontal; Prf, prefrontal; Pt, pterygoid; Q, quadrate; Qj, quadratojugal; Sq, squamosal; St, supratemporal; T, tabular.
- Temporal emargination bordered by quadratojugal and squamosal (Fig. 2C, D). The temporal emargination supports a tympanum in turtles, and the presence of a slender stapes in several anapsids suggests that a tympanum appeared fairly early in this group. There is no temporal emargination in romeriids, mesosaurs, and synapsids (Fig. 2A, B).
- Jaw articulation anterior to occiput (Fig. 1C, D). The jaw articulation of romeriids, mesosaurs, synapsids, and diadectomorphs is at the level of the occiput (Fig. 1A, B).
- Edentulous ectopterygoid. The ectopterygoid bears a shagreen of small denticles in romeriids, early synapsids, and diadectomorphs.
- Stapedial dorsal process unossified. The stapes of most early romeriids, mesosaurs, synapsids, and diadectomorphs has an ossified dorsal process.
- Sacral ribs with narrow distal contact. The sacral ribs of romeriids, mesosaurs, and synapsids are broad distally and a broad contact between successive ribs leaves only a small gap. The sacral ribs of anapsids are more slender and contact each other distally.
- Iliac blade dorsally expanded. The iliac blade of romeriids, mesosaurs, and synapsids has a long, low posterodorsal process and lacks an anterior expansion. The iliac blade of anapsids has a dorsal expansion directly above the acetabulum, a short anterior process, and it is short posteriorly.
Figure 3. Skulls of early amniotes in dorsal view. A, Cotylorhynchus, a Lower Permian synapsid; B, Captorhinus, a Lower Permian romeriid; C, Procolophon, a Triassic anapsid; D, Proganochelys, the oldest known turtle (Upper Triassic). Redrawn from A, Laurin & Reisz (1995); B, Heaton (1979); C, Carroll & Lindsay (1985); and D, Gaffney (1990). Abbreviations: Bc, braincase; F, frontal; J, jugal; L, lacrimal; M, maxilla; N, nasal; P, parietal; Pa, palatal; Pm, premaxilla; Po, postorbital; Pof, postfrontal; Pp, postparietal; Prf, prefrontal; Qj, quadratojugal; Sq, squamosal; St, supratemporal; T, tabular.
Procolophonia is defined as "The most recent common ancestor of pareiasaurs, procolophonids, and testudines (Chelonia), and all its descendants" (Laurin & Reisz, 1995). It is diagnosed by several autapomorphies, including the following:
- Pineal foramen close to fronto-parietal suture (Fig. 3C). The pineal foramen of most other amniotes is located close to the mid-length of the interparietal suture, or slightly anterior to this (Fig. 3A, B).
- Tabular absent (Fig. 3C, D). Most other amniotes have a tabular (Fig. 3A), but this bone has been lost convergently in captorhinids (Fig. 3B).
- Cranio-quadrate space large. In most other amniotes, the cranio-quadrate space (the space between the braincase and the quadrate ramus of the pterygoid) is narrow and the paroccipital process and the quadrate ramus of the pterygoid converge posterolaterally (Fig. 1A, B). In procolophonoids, pareiasaurs, and turtles, the cranio-quadrate space is wide and the paroccipital process is parallel to the quadrate ramus of the pterygoid (Fig. 1C, D).
- Pterygoid palatal ramus not reaching level of internal naris (Fig. 1C, D). The palatal ramus of millerettids, romeriids, mesosaurs, and early synapsids extends anteriorly medial to the internal naris (Fig. 1A, B).
- Transverse flange of pterygoid directed anterolaterally (Fig. 1C, D). The transverse flange of other amniotes extends transversely or posterolaterally from the area of the basicranial articulation (Fig. 1A, B).
- Cultriform process short (Fig. 1C, D). The cultriform process of other amniotes is long (Fig. 1A, B).
- Supraoccipital plate reduced to a sagittal pillar.
- Paroccipital process antero-posteriorly expanded. The paroccipital process of most other amniotes is a broad, thin, vertical flange.
- Quadrate condyle articular surfaces nearly flat and antero-posteriorly short. In other amniotes, the articular surfaces of the quadrate are strongly convex and short.
- Astragalus and calcaneum (two ankle bones) sutured or fused to each other.
- Loss of the fifth distal tarsal (an ankle bone).
Romeriida is defined as "extant saurians, and all other taxa that are more closely related to them than they are to anapsids" (Gauthier et al., 1988). Romeriida possesses the following autapomorphies:
- Postorbital separated from supratemporal (Fig. 3B). The postorbital contacts the supratemporal in most anapsids (Fig. 3C), in mesosaurs, in early synapsids (Fig. 3A), and in diadectomorphs.
- Supratemporal small (Fig. 3B). The supratemporal is large in most other groups of amniotes (Fig. 3A, C).
- Caniniform maxillary tooth present. Romeriids have a large anterior maxillary tooth (Fig. 2B). Most other early amniotes have a relatively homodont dentition and no anterior maxillary tooth is much larger than the other teeth (Fig. 2A, C).
- Quadrate anterior process short (Fig. 1B). The anterior process of the quadrate extends anteriorly along more than 55% of the quadrate ramus of the pterygoid in most other amniotes (Fig. 1A, D).
"Protorothyridids" and diapsids share the following synapomorphies:
- Anterior pleurocentra keeled ventrally. The anterior pleurocentra of other amniotes is rounded ventrally.
- Carpus and tarsus long and slender.
- Metapodials overlapping. The proximal heads of the metapodials of other amniotes barely contact each other. The metapodials of "protorothyridids" and diapsids are expanded proximally and overlap the metapodial lateral to them.
We thank Miss Diane Scott for scanning the figures of amniote skulls, and Ms. Patricia Lai for proof-reading this page.