




Passeriformes
Perching Birds
Scott V. Edwards



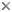
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Passeriformes is the largest and most diverse commonly recognized clade of birds. The Passeriformes (or ‘passerine’ birds) are synonymous with what are commonly known as "perching birds"; it also contains within it a major radiation commonly known as songbirds (oscine Passerines or Passeri). Of the 10,000 or so extant species of birds, over half (~5,300) are perching birds. Perching birds have a worldwide distribution, with representatives on all continents except Antarctica, and reaching their greatest diversity in the tropics. Body sizes of passerines vary from about 1.4 kg in northern populations of Ravens (Corvus corax) to just a few grams. Perching birds include some of the most colorful and mysterious of all birds, such as birds of paradise from New Guinea and the bright orange Cock of the Rock from tropical South America. Because of their high diversity, generally small body size and relative ease of observation, collection and field study, perching birds have historically attracted the attention of a wide range of descriptive and experimental biologists, including systematists, behavioral ecologists, and evolutionary biologists.
Monophyly and Sister Group
Historically, it is generally agreed that the Passeriformes constitute a monophyletic group. Raikow (1982) established this monophyly in an explicitly phylogenetic context. He noted that Passeriformes possess a suite of distinguishing characteristics, including a unique sperm morphology, a distinctive morphology to of the bony palate, a simple yet functionally diverse foot with three toes forward and one (the hallux) oriented backwards, and a distinctive fore- (wing) and hindlimb musculature. There are few if any species which pose problems for avian systematists as to whether they are or are not passerines. Most of the controversy lies in relationships within the clade.
The sister group of the Passeriformes is not so much hotly contested as it is poorly resolved by existing data sets. Traditionally, Passeriformes have been considered closely related to a large group known as the “higher non-Passerines”. These include a number of clades such as cuckoos (Cuculiformes), hornbills, kingfishers and related lineages (Coraciiformes), and woodpeckers and relatives (Piciformes). Many of these groups possess a zygodactyl foot, a condition in which two toes point forward and two point backward. The sister relationship of Passeriformes to woodpeckers, the hornbill group and allies is reflected in Joel Cracraft’s phylogenetic hypothesis for major groups of birds based on cladistic interpretation of morphological and molecular characters (Cracraft, 1988). However, in the other major classification bearing on the relationships of perching birds, that based on DNA-DNA hybridization, Passeriformes appear as the sister group to a large, diverse group containing pigeons and doves (Columbiformes), cranes and rails (Gruiformes) and storks (Ciconiiformes)! These latter three groups share few obvious morphological characteristics with Passeriformes. However, the DNA hybridization tree links Passeriformes with these groups at a very deep level in the tree, rendering this result tenuous. Clearly, more work on the sister-group relationship of Passeriformes is needed, since this relationship will be the basis of any study seeking to identify whether or not Passeriformes are a particularly diverse group (e.g., Nee et al. 1992).
Major Groups of Passeriformes
The basal lineages of passeriformes have for over a century included two major clades the oscines and suboscines. Suboscines (Tyranni) are a largely tropical group of about 1,000 species that reaches its greatest diversity in the South America; most suboscines are thought to sing “innate” songs. Oscines (Passeri) include about 4,000 species and are what many laypersons refer to as “songbirds”; they are worldwide in distribution and are distinguished from suboscines by a complex voice box (syrinx) and song learning capacity. The German anatomist Müller first drew attention to the syringeal differences (Müller, 1878), and since then both morphological (Ames, 1971) and molecular (Edwards et al., 1991; Sibley and Ahlquist, 1990) analyses have confirmed this basic division of perching birds. However, there are a few species, such as the two lyrebirds in Australia (Espinosa and Cracraft, 1994; Feduccia and Olson, 1982), and the diminutive New Zealand wrens (Acanthisitidae; Raikow, 1987), that evade easy placement into either of these groups . The lyrebirds, for example, possess extremely complex vocalizations and molecular affinities to oscines, but other morphological traits and biogeographically some consider them more closely related to suboscines.
It is worth mentioning here one more division within the oscine passerine birds. Prior to the application of molecular data to questions of higher level passerine phylogeny, there was very poor if any resolution of the major clades. However, DNA-DNA hybridization comparisons (Sibley and Ahlquist, 1990) have shed enormous light on this issue and delineated a major division within the oscines. These studies suggest that the oscines consist of two major groups, the Corvida, consisting of crow-like species and others that originated in the Australo-Papuan region, and the Passerida, consisting of sparrow- and thrush- like species, and which are found mainly in Eurasia, Africa and the Americas. This molecular division of oscines into Passerida and Corvida is corroborated by at least one morphological character, namely the condition of the tricipital fossa at the head of the humerus. The Corvida possess the primitive state for this trait, a single fossa that is highly pneumatic (also found in suboscines), while the Passerida possess a double fossa that is less pneumatic. In older classifications of oscines (Wetmore, 1960), the condition of the fossa appeared to be scattered across the families. Whole families of birds that used to be interpreted as close relatives spread across Australia and Eurasia now are thought to be spectacular examples of convergence. Australia and New Guinea have their own “robins”, “wrens” and “treecreepers”, but they are not at all closely related to lineages of the same name found elsewhere.
Diversity of Passeriformes
Species diversity: The tradition of recognizing perching birds (Passeriformes) as the most diverse and rapidly radiating clade has been questioned because there are few obvious “key innovations” that should cause systematists to recognize Passeriformes over any other arbitrarily larger or smaller monophyletic group within birds (Raikow, 1986). One point that has been missed in debates on this issue is that the branch leading to the songbirds (oscines), a group comprising 80% of extant perching birds, is the longest internal branch on the DNA hybridization tree produced by Sibley and Ahlquist (1990). This branch has also been one of the few to be well resolved in applications of mtDNA sequences to higher level questions in birds, presumably because it is long. Given the large number of clades that will require names under phylogenetic taxonomy, perhaps the length of branches leading to particular clades should be one criterion whereby systematists decide which of the many clades to name.
Behavioral diversity: It is extremely difficult to generalize about any of the behaviors or nesting habits of passerines, because as a group they are so diverse. Perching birds exhibit a bewildering array of plumages and colors derived from diverse keratin structures as well as ingested pigments, such as carotenoids (Gray, 1996). Many passerines, such as some Old World Flycatchers (Muscicapidae) and African Widowbirds (Viduinae) have extremely long tail feathers or highly modified plumes (Birds of Paradise: Paradisaeidae) used in courtship displays,. Several groups such as the Wattlebirds of New Zealand (Callaeidae) and Honeyeaters (Meliphagidae) have fleshy, bright blue, red or yellow wattles on the face and neck. Perching birds build their nests generally out of sticks or grass on the ground, in trees, and in the case of Dippers (Cinclidae) in the banks of fast-flowing rivers. Many passerines migrate from their nesting grounds in the Nearctic and Palearctic to more equatorial regions, or from southern temperate regions north to the tropics. Parental care by both sexes is common in passerines, although some highly dimorphic and predominantly lekking groups, such as manakins (Prum, 1994) and birds of paradise (Diamond, 1986), females alone provide for young and build the nest. Cooperative breeding, in which young birds delay breeding and assist other individuals (often parents) in raising young and defending the territory, is common in several passerine groups, such as Australian fairy wrens (Maluridae) and New World Jays (Corvidae; Brown, 1987; Edwards and Naeem, 1993). Some of the most elaborate singers in the bird world are passerines (Kroodsma and Miller, 1996). Some passerine birds are poisonous to the touch and are avoided as prey for indigenous peoples (Dumbacher et al., 1992).
Origin and biogeography of passerines: The temporal and geographic origin of passerine birds is obscure. Traditionally the group was thought to have originated in the Tertiary, at about the same time as extant orders of mammals. Some recent workers favor a later, Eocene origin (Feduccia, 1995; Wilson, 1989), but the DNA -DNA hybridization data again favors an earlier origin (Sibley and Ahlquist, 1990). Recently some of the oldest oscine fossils have been uncovered in Queensland, Australia (Boles, 1995); this and other paleobiogeographical data suggest that passerines may have in fact originated in the Southern hemisphere (Olson, 1989).
References
Ames, P. L. (1971). The morphology of the syrinx in passerine birds. Bulletin of the Peabody Museum of Natural History 37, 1-94.
Boles, W. E. (1995). The world's oldest songbird. Nature 374, 21-22.
Brown, J. L. (1987). Helping and Communal Breeding in Birds. In Monographs in Behavioral Ecolog. Princeton, N.J.: Princeton University Press.
Cracraft, J. (1988). The major clades of birds. In The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds, vol. 35A (ed. M.J.Benton), pp. 339-361. Oxford: Clarendon Press.
Diamond, J. (1986). Biology of birds of paradise and bowerbirds. Annual Review of Ecology and Systematics 17, 17-37.
Dumbacher, J. P., Beehler, B. M., Spande, T. F., Garraffo, H. M. and Daly, J. W. (1992). Homobatrachotoxin in the genus Pitohui : chemical defence in birds? Science 258, 799-801.
Edwards, S V, Arctander, Wilson and A C. (1991). Mitochondrial resolution of a deep branch in the genealogical tree for perching birds. Proceedings of the Royal Society of London series B 247, 99-107.
Edwards, S. V. and Naeem, S. (1993). The phylogenetic component of cooperative breeding in perching birds. The American Naturalist 141, 754-789.
Espinosa, A. and Cracraft, J. (1994). Molecular phylogeny of the lyrebird. Journal f?r Ornithologie 135, 29.
Feduccia, A. (1995). Explosive radiation in Tertiary birds and mammals. Science 267, 637-638.
Feduccia, A. and Olson, S. L. (1982). Morphological similarities between the Menurae and the Rhinocryptidae, relict passerine birds of the Southern hemisphere. Smithsonian Contributions to Zoology 366, 1-22.
Gray, D. A. (1996). Carotenoids and sexual dichromatism in North American passerine birds. The American Naturalist 148, 453-480.
Kroodsma, D. E. and Miller, E. H. (1996). Ecology and Evolution of Acoustic Communication in Birds. Ithaca, NY: Cornell University Press.
M?ller, J. P. (1878). On certain variations in the vocal organs of the Passeres that have hitherto escaped notice. London: Macmillan.
Nee, S., Mooers, A., and Harvey, P. H. (1992). Tempo and mode of evolution revealed from molecular phylogenies. Proceedings of the National Academy of Sciences (USA) 89, 8322-8326.
Olson, S. L. (1989). Aspects of global avifaunal dynamics during the Cenozoic. In Proceedings of the XIX International Ornithological Congress, vol. 29, pp. 2023-2029. Christchurch, New Zealand.
Prum, R. O. (1994). Phylogenetic analysis of alternative social behavior in the manakins (Aves: Pipridae). Evolution 48, 1657-1665.
Raikow, R. J. (1982). Monophyly of the Passeriformes: test of a phylogenetic hypothesis. Auk 99, 431-455.
Raikow, R. J. (1986). Why are there so many kinds of passerine birds? Systematic Zoology 35, 255-259.
Raikow, R. J. (1987). Hindlimb myology and evolution of the Old World suboscine passerine birds. Ornithological Monographs 41, 1-81.
Sibley, C. G. and Ahlquist, J. E. (1990). Phylogeny and Classification of Birds. New Haven, CT: Yale University Press.
Wetmore, A. (1960). A Classification for the Birds of the World. Smithsonian Miscillaneous Collections 39, 1-37.
Wilson, A. C. (1989). Time scale for bird evolution. Proceedings of the XIX International Ornithological Congress 19, 1912-1917.
Title Illustrations

| Scientific Name | Carpodacus mexicanus |
|---|---|
| Comments | A House Finch (Passerida, Fringillidae), a common North American songbird that was introduced to the Eastern U.S. in the mid-1940s. |
| Specimen Condition | Live Specimen |
| Sex | Male |
| Copyright | © 1997 Geoff Hill |

| Scientific Name | Agelaius phoeniceus |
|---|---|
| Comments | Red-winged Blackbirds. Males of this polygynous species defend territories and are a classic example of resource-base polygyny in birds. In this mating system, females are predicted to achieve greater reproductive success with males with one or more females on high quality territories than with males with no females on poor territories. |
| Specimen Condition | Live Specimen |
| Sex | Male |
| Copyright | © 1997 Gordon Orians |
About This Page
I willingly invite more web-facile passerine enthusiasts to contribute suggestions and photographs that will improve this page. I thank David Maddison for formatting the page.
Scott V. Edwards
Department of Organismic and Evolutionary Biology
Harvard University
26 Oxford Street
Cambridge, MA 02138
Correspondence regarding this page should be directed to Scott V. Edwards at
Page copyright © 1997 Scott V. Edwards
- First online 18 July 1997
Citing this page:
Edwards, Scott V. 1997. Passeriformes. Perching Birds. Version 18 July 1997 (under construction). http://tolweb.org/Passeriformes/15868/1997.07.18 in The Tree of Life Web Project, http://tolweb.org/
